Thesis for the degree Doctor of Philosophy Submitted to the Scientific Council of the Weizmann Institute of Science Rehovot, Israel עבודת גמר )תזה( לת
|
|
|
- אבראהים אדלר
- לפני4 שנים
- צפיות:
תמליל

1 Thesis for the degree Doctor of Philosophy Submitted to the Scientific Council of the Weizmann Institute of Science Rehovot, Israel עבודת גמר )תזה( לתואר דוקטור לפילוסופיה מוגשת למועצה המדעית של מכון ויצמן למדע רחובות, ישראל By Boaz Gildor מאת בועז גילדור מהתפתחות לביצוע: איחוי תאי שריר בזבוב הפירות From development to execution: Myoblast fusion in Drosophila Advisors: Prof. Ben-Zion Shilo Dr. Eyal Schejter מנחים : פרופ' בן-ציון שילה ד"ר אייל שכטר July אלול תשע"א
2 תודות כמיטב המסורת אני מעוניין להקדיש את העבודה המוצגת כאן לאנשים הרבים שעזרו ליצירתה: למנחים שלי, בני ואייל, שליוו אותי כמעט שש שנים, עם הרבה קשב והמון תמיכה. לבני, שלימד אותי רבות על מוטיבציה, יעילות, הכוונה ומנהיגות. בלעדייך הייתי הולך לאיבוד בדרך מזמן. ולאייל, שלימד אותי על אהבת המדע, כבוד לסובבים אותי, מתינות וצניעות. בזכותך אני יודע היום שיש הרבה יותר מדרך אחת לעשות מדע טוב. לשרי כרמון, שתמיד דאגה לכך שארגיש בבית - מוגן ואהוב. לחבריי למעבדה, שליוו אותי לאורך השנים וקיבלו אותי בזרועות פתוחות: אפרת אסא-קוניק, מירב אריאלי, דני בן-צבי, ארז גרון, מיכל השכל-איטח )הסטודנטית הראשונה שהדרכתי וכנראה גם הטובה ביותר שאני אי פעם אדריך(, שאול יוגב )חבר יקר, מדען מבריק ואדם למופת(, יעל כהן, ורד כץ, לאה סננס, ראדה מסארוואה )שהיוותה לא רק את סוללת הדרך לעבודה זאת אלא גם מודל לחריצות(, שי רוטקופף, טל רוסו )שעזרה לי אין ספור פעמים בעבודה המדעית והעלתה לי חיוך על הפנים יום-יום(, ארקדי שוורץ, גלית שוחט, תמר שפירא-כהן. תודה מיוחדת לפריאנקה מוקרג'י, מבנגלור שבהודו, שלימדתה אותי את הכלים הדרושים לחלק ניכר מהעבודה. אך כמובן שלא רק בזכות אנשי מכון ויצמן הפרויקט יצא לאור. אני רוצה להודות מכל הלב לאנשים שמילאו את החלק האחר והאישי,של חיי באהבה ללא גבולות ובתמיכה מתמדת: להוריי, חוה ויעקב, שתמיד היו לי, ולתמיד יהיו לי לעיר מקלט. לאחיי, גיא ועופר, מקור השראה ודוגמא לחיים טובים, מהנים, יעילים ומלאי שליחות. לבן זוגי היקר, עמרי, שמלווה אותי לכל אורך הדרך - זאת והעתידות לבוא. לכולכם אני רוצה לומר- תודה. נ.ב- וכמובן לחתול שלי, מיו, שדאג לכך שהרעיונות הכי טובים הגיעו כשהוא יושב בחיקי. 2
3 Abstract In most organisms muscles are multi-nucleated cells, that are generated by cellcell fusion. In this thesis, I will present work utilizing the Drosophila myoblast fusion system as an experimental tool to investigate this remarkable part of myogenesis. Myoblast fusion in Drosophila is relatively well described, and many necessary factors have been identified. This enabled us to ask advanced questions regarding required cellular remodeling involved in the process. The fly life cycle offers to us a powerful tool for comparison between two distinct biological processes: embryonic and adult myoblast fusion. Both processes occur in the same organism but each presents its own requirement from the fusion machinery. The results from this project are presented here in three chapters. 1. Examining the roles of the actin nucleation factors WASp and SCAR/WAVE during embryonic myoblast fusion. Both factors drive the nucleation of branched actin filaments through activation of Arp2/3, and we observed that both are essential for embryonic myoblast fusion. Such a requirement is surprising, given the similar outcome of their activation. However, careful phenotypic examination suggests that they function in a sequential fashion. SCAR activity is required prior to the formation of pores in the membranes of fusing cells, while WASp promotes expansion of nascent pores and completion of the fusion process. 2. The roles of WASp and SCAR/WAVE are not restricted to embryonic fusion. WASp is described as the first bona-fide mutant to disrupt adult fusion during metamorphosis, and we have observed that the roles of the SCAR/WAVE in the process are also preserved. While both factors are required for adult fusion, the creation of Arp2/3-dependent branched actin foci is mediated by a third, unknown, actin nucleator. 3. Finally, we utilize RNAi technology in order to examine, at the pupal stage, genes that were shown to be required for myogenesis and myoblast fusion in the embryo. We show that these genes are also required for muscle formation in the pupa. More specifically, that the cell adhesion protein Dumbfounded (Duf) is required for attraction of migrating myoblasts to the myotubes, as well as for the initial contact between the myotube and the myoblasts. When migrating as a swarm, the myoblasts are maintained in the transit-amplifying state by continuous Notch activation, where each myoblast 3
4 provides the source of ligand, Delta (Dl), to its neighbors by fine-tuning the levels of ligand and receptor it displays. This activation represses expression of Sticks and stones (Sns). Only upon reaching the vicinity of the myotubes does Notch signaling decay, leading to terminal differentiation of the myoblasts and triggering the onset of fusion by induction of Sns expression. Generally, we found that the overall usage of proteins during embryonic and pupal myoblast fusion is similar, but modifications in the program allow to adjust it to the particular biological setting of each stage. 4
5 תקציר ברוב האורגניזמים שרירים הם תאים רבי-גרעין, הנוצרים בתהליך של איחוי בין-תאי. בתזה המוצגת כאן, אני אציג עבודה המנצלת את תהליך איחוי תאי השריר בזבוב הפירות, תסיסנית המחקר, ככלי ניסויי לחקר התהליך המופלא הזה הנדרש במהלך יצירת השרירים. איחוי תאי שריר בזבוב הפירות תואר בצורה מעמיקה במחקרים קודמים, ורבים מהגורמים הנחוצים לו נחקרו. כלי זה איפשר לנו לשאול לשאול שאלות מורכבות בנוגע לתקשורת הבין-תאית והשינויים התאיים הדרושים לביצוע התהליך. מחזור החיים של הזבוב העמיד בפנינו כלי יעיל להשוואה בין שני תהליכים שונים מבחינה ביולגית: איחוי תרי שריר בזמן התתפתחות העוברית ואיחוי תאי שריר בזמן ההתגלמות. שני התהליכים מתרחשים באותו האורגניזם אבל מציבים דרישות ביולוגיות שונות ממנגנון האיחוי. התוצאות מפרויקט זה מוצגות כאן בשלושה פרקים: 1. בחינה של תפקידם של פקטורי גירעון האקטין, WASp ו-,SCAR/WAVE בזמן איחוי תאי שריר בעובר. שני הפקטורים גורמים לגירעון של אקטין, ודוחפים לפולימריזציה של סיבי אקטין מסועפים דרך הפעלת קומפלקס ה-,Arp2/3 ואנו גילינו ששניהם דרושים לצורך ביצוע איחוי תאי השריר בעובר. דרישה זאת היא מפתיעה, לאור התוצאה הדומה של הפעלתם של שני הפקטורים. אך בחינה מעמיקה של הפגיעה בכל אחד משני הפקטורים חשפה שהם פועלים בצורה עוקבת. הפעילות של SCAR דרושה לפני הופעת חורים בקרומי שני התאים המתאחים, בעוד שפעילותו של WASp דוחפת את הרחבתם של החורים ונחוצה לסיום התהליך בהצלחה. 2. פעילותם של WASp ו- SCAR/WAVE איננה מוגבלת לאיחוי תאי שריר בזמן התתפתחות העוברית. WASp תואר כמוטנט המובהק הראשון הפגוע בתהליך איחוי תאי שריר בזמן ההתגלמות, ואנו גילינו שגם תפקידו של קומפלקס ה- SCAR/WAVE בתהיך שמור. בעוד ששוב, שני הפקטורים דרושים לביצוע האיחוי, יצירת סיבי אקטין מסועפים ע"י קומפלקס ה- Arp2/3 מתווכת ע"י פקטור שלישי, שאיננו ידוע. 3. לבסוף, הסתייענו בשימוש בטכנולוגיית RNAi כדי לבחון בזמן המטמורפוזה, את תפקידם של גנים שהוכחו כחיוניים לתהליך התפתחות השרירים ואיחויים בעובר. התוצאות שאנו מציגים מראות שגנים אלה דרושים גם להתפתחות שרירי הבוגר בזמן ההתגלמות. ספציפית, חלבון האדהזיה (Duf) Dumbfounded דרוש לצורך משיכת תאי השריר לשרירים הנוצרים ובנוסף לצורך המגע הראשוני בין תאי השריר לשרירים הגדלים. בזמן שתאי השריר נודדים כנחיל, הם מוחזקים במצב התמיינותי של "התחלקות זמנית" ע"י חשיפה לאות.Notch כל אחד מתאי השריר בנחיל מבטא את מקור הליגנד לאות, (Dl),Delta ומציגו בפני תאי השריר השכנים שלו בנחיל, בעזרת כיוונון עדין של רמות הליגנד והרצפטור שהוא מציג. הפעלה זאת 5
6 של מסלול Notch מונעת את ביטוי הגן (Sns).Sticks and stone רק לאחר שהתאי השריר הנודדים מגיעים בסמוך לסיבי השריר, אות ה- Notch דועך, דבר המוביל להתמיינות הסופית של תאי השריר וגורר איחוי של תאי השריר בעזרת השרייה של ביטוי של.Sns בכלליות, אנו מוצאים שחלבונים דומים נדרשים לצורך איחוי תאי שריר בעובר ובבוגר, אך שינויים בתוכנית הגנטית של תהליך האיחוי מאפשרים להתאים אותו לסביבה הביולוגית היחודית בכל שלב התפתחותי. 6
7 List of abbreviations AP- adult precursor Blow- blown fuse DC- dendritic cell Dl- Delta DLM- dorsal longitudinal muscles Duf- dumbfounded DVM- dorsal ventral muscles EM- electron microscope FC- founder cell FCM- fusion competent myoblast GEF- guanine exchange factor Hbs- Hibris IFM- indirect flight muscles Mbc- myoblast city N- Notch NPF- nucleation promoting factor Rst- roughest SCAR- suppressor of cyclic AMP receptor Ser- Serrate Sing- singles bar Sns- sticks and stones SOP- sensory organ precursor cell TF- transcription factor WASp- Wiskott Aldrich syndrome protein WIP- WASp interacting protein 7
8 Table of contents 1. General introduction Myogenesis in Drosophila Embryonic Drosophila Myogenesis Embryonic Myoblast Fusion in Drosophila Actin Polymerization by Arp2/3 and its Activators Metamorphosis and the Formation of the Adult Musculature Notch signaling pathway Cis-inhibition of Notch signaling The SCAR and WASp nucleation promoting factors act sequentially to mediate Drosophila myoblast fusion Absrtact Introduction Results and discussion Multiple roles for the SCAR complex during embryonic myogenesis Distinct, sequential roles for SCAR and Wsp during myoblast fusion...28 The formation of actin foci at fusion sites is not mediated by SCAR and Wsp NPF activity Methods Acknowledgements Supplemental information The Actin Nucleator WASp is Required for Myoblast Fusion During Adult Drosophila Myogenesis Introduction Results The Arp2/3 nucleation-promoting factor SCAR is required for adult myoblast fusion Arp2/3, but neither of its major NPFs, mediates formation of fusion- associated actin foci
9 3.4. Discussion Involvement of the Arp2/3 nucleation system in formation of fusionassociated actin foci Fusion competence is repressed in migrating Drosophila myoblasts by Notch signaling Introduction Results Duf provides an attractive cue to migrating imaginal myoblasts Duf is required for fusion Sns/Hbs function mediates establishment of myoblast-myotube fusion sites Sns is expressed in terminally differentiated adult myoblasts...65 Notch signaling maintains the transit-amplifying state Delta-Notch signaling between myoblasts prevents premature differentiation A fine balance between transmitting and receiving Notch signals in myoblasts Discussion Methods Supplemental information General discussion Common mechanism utilized repeatedly for myoblast fusion Constancy and modulations of the fusion machinery Future prospects for the analysis of adult Drosophila myogeneis WASp and SCAR- two, but not the same Controversy regarding the roles of SCAR and WASp in myoblast fusion Notch- an antagonist of adult myogenesis Cis inhibition- dogma and exceptions Literature List of publications Declaration
10 1. Introduction 1.1. Myogenesis in Drosophila The life cycle of Drosophila includes two major and distinct phases of development- embryogenesis, which leads to formation of a larva; and metamorphosis, which includes destruction of larval tissues and generation of the adult fly. Each of these two phases of development includes formation of different sets of somatic muscles. During embryonic development, myogenesis generates muscle elements that mediate the relatively simple behavioral repertoire of the larva, while during pupal metamorphosis, a diverse pattern of muscle fibers is assembled, and these facilitate the more elaborate behavioral patterns of the adult fly [1, 2]. In this thesis, I will present work concerning different aspects of the development of the multi-nucleated somatic musculature in the fruit fly. My work is mainly focused on the execution of fusion between muscle cells in the embryo and adult. I will start by presenting background knowledge on the development of the somatic muscles throughout the Drosophila life cycle Embryonic Drosophila myogenesis During early Drosophila embryonic development, visceral, heart and somatic muscles establish their identity. All muscles originate from a mesodermal origin. The somatic muscles are positioned underneath the epidermis in a segmentally repeated pattern, ~30 per hemisegment and ~600 in total [3, 4]. The patterning of somatic muscles includes several processes: myoblast specification, fusion, migration and attachment. One of the first events to occur is the specification of two cell populations out of a field of equivalent myogenic cells. This is achieved through a process called- lateral inhibition. Initially, all cells express the Notch receptor and the ligand Delta (for more on Notch signaling please see 1.6.). Fluctuations in the levels of gene expression eventually cause a single cell from a group of about 7 cells to be specified and become dedicated to Delta expression, at the expense of expressing Notch and receiving the signal. This cell will now signal to all its neighbors to up-regulate Notch and repress Delta. These Delta expressing, signal sending cells will become the founder cells (FCs), and all the Notch 10
![receiving cells will become fusion competent myoblasts (FCMs) [5]. The founder cells are responsible for the uniqueness of each muscle fiber identity.](/docs-images/104/165733799/images/11-0.jpg "The FCMs, in contrast, adopt a more naïve fate, and will gain the ability to fuse to any founder cell.")
11 receiving cells will become fusion competent myoblasts (FCMs) [5]. The founder cells are responsible for the uniqueness of each muscle fiber identity. The FCMs, in contrast, adopt a more naïve fate, and will gain the ability to fuse to any founder cell. Each founder cell will now undergo an asymmetric division and will form a single founder cell and a single adult precursor (AP). By asymmetric distribution of the Notch repressor, Numb, the AP will become Notch positive and will not participate in the embryonic fusion process. The APs will continue to express early mesodermal markers such as Twist, and will migrate to specific locations in the embryo. These cells will have a pivotal role in the formation of the adult musculature[6]. Following the differentiation of these cell types, each founder cell fuses with a stereotypic number of FCMs to form a particular muscle fiber. Afterwards, the growing fibers migrate towards specific epidermal attachment sites and form adhesion junctions with their target cells [7, 8]. Scheme 1. Schematic representation of the different steps of Drosophila myogenesis, adapted from [8]. Progenitors (P, orange) are singled out from a field of myogenic competent cells (blue) and give rise to two founder cells or an adult precursor (AP, grey) after one asymmetrical division. The founders fuse with the FCMs (green) to form multinucleated fibers, which migrate and eventually attach to their specific targets at the epidermis. 11
12 1.3. Embryonic Myoblast Fusion in Drosophila Myoblast fusion, which is at the focus of this study, is a multistep process. After the myoblasts become specified as either FCs or FCMs, these cells must identify and adhere to their fusion partners. Recognition is specific and directional, in that fusion does not occur between two myoblasts of the same type. Genetic studies in Drosophila embryos have identified a number of key players in the fusion process. These studies begin to define the genes and steps necessary for the recognition and adhesion between myoblasts, and the fusion of the two lipid bilayers into one. In a detailed morphological analysis of fusing myoblasts at the level of the electron-microscope (EM) [9], a series of intracellular events that accompany fusion was described (Scheme 3). The first obvious change is the accumulation of clusters of electron dense vesicles on the apposed plasma membranes of two associated myoblasts. These vesicles, which have not been observed in other cell types, will eventually align with each other across the intervening membranes to form paired vesicles. After alignment, the vesicles are thought to resolve into electron-dense plaques, which span the plasma membranes. Multiple pores are then observed in the fusing membranes, usually adjacent to the electron-dense plaques. Theses pores expand in size, until complete dissolution of the membrane is achieved [9]. A variety of mutations that affect the fusion process have been reported, and for many of them the exact step in which they are arrested has been observed by electron microscopy. Four members of the immunoglobin superfamily are known to be involved in embryonic myobaslt fusion: dumbfounded/kin of irrec (duf/kirre)[10], roughest (rst) [11], sticks and stones (sns)[12, 13] and hibris (hbs)[14, 15]. Duf is expressed exclusively in founder cells and fusing myotubes while rst, which is functionally redundant of duf, has a less restricted pattern of expression and it can also be seen in some myoblasts. Sns is expressed in all myoblasts and is quickly downregulated once these fuse to myotubes. It was recently proven that although hbs is only homologous to sns in the extracellular domain, it functions in a redundant fashion to sns, and in order to receive the full phenotype both need to be mutated [15]. Biochemical studies revealed that the extarcellular domains of Duf and SNS can bind to each other [12, 13]. 12
13 It appears that this interaction is crucial both for the chemotaxis of the migrating FCMs towards the founders, and for the adhesion of the two cell types [13]. When tested in cell culture, Duf-expressing cells secrete the truncated extracellualr domain of the protein to the medium [10]. In embryos mutant for sns or duf the FCMs fail to attach to the founders, so that the fusion is halted at the very beginning of the process. An interesting observation is that FCMs mutant for sns fail to form lamellopodia, which will direct their migration to the founder, while in embryos mutant for duf the lamellopodia are formed, but at random directions [10]. The intracellular events that occur during fusion have also been associated with distinct genes. Several other genes have emerged from screens aimed to identify genes required for embryonic myoblast fusion. One of these is myoblast city (mbc), a non-typical guanine exchange factor (GEF) that when in a complex with ELMO, was shown to activate rac, another crucial gene in myoblast fusion [16-22]. Several other genes were first described for their role in myoblast fusion such as the PH domain protein blown fuse (blow) [9, 20] or MARVEL domain protein singles bar (sing) [23]. Although it is known that they function in fusion at a distinct phase, their molecular role or the pathway they take part in are still not understood. Scheme 3 summarizes the steps at which the fusion arrests, in the corresponding mutants. Scheme 3. Myoblast fusion is a multistep process. (i) A fusion-competent cell (green) extends filopodia towards a binucleated muscle precursor (brown). (ii) The fusion-competent cell recognizes and attaches to 13
14 the muscle precursor. (iii) Paired vesicles with electron-dense margins (prefusion complexes) form along the apposed membranes. (iv) An electron-dense plaque forms along the membranes. (v) The cells align along their entire long axes. (vi) The apposed membranes break down, accompanied by the formation of fusion pores. (vii) A multinucleated myotube is formed. The genes illustrated in blue have been shown to function at different stages of myoblast fusion. Adapted from [24] Actin Polymerization by Arp2/3 and its Activators Cell processes involving movement and changes in shape rely on cycles of actin polymerization and depolymerization. Actin filament nucleation is energetically unfavourable; therefore, nucleation factors are required to initiate assembly of any actinbased structure. Three major classes of nucleation factors have been characterized (Arp2/3 complex, Formin proteins and Spire [25]), although additional nucleators are constantly emerging [26]. Most notably, the Arp2/3 complex is responsible for nucleating de novo actin filaments at the sides of existing filaments, resulting in a branched array that produces forces, which can be applied to various cellular factors, such as pushing the leading edge of cells forward. Arp2/3 alone does not stimulate actin filament production. Rather, when cells are stimulated it is activated by a range of accessory proteins [27]. The WASP family of proteins is the best understood class of Arp2/3 activators; they coordinate actin reorganization by coupling signaling molecules, including Cdc42-GTPases, to activation of the Arp2/3 complex [28]. The regulatory mechanisms of the two main groups of WASP family proteins WASPs and SCAR/WAVEs are quite different. WASp is regulated primarily by autoinhibition that is relieved by direct binding of the GTPase Cdc42 to its GTPase-binding domain [29]. However, understanding how SCAR/WAVEs are regulated has proved more difficult, as they have no GTPase-binding domain and their activity profile at the purified state is still under controversy. A paper linking WAVE1 to a complex of four other proteins has stimulated exciting progress in this area. Inactive WAVE1 (SCAR in Drosphila) was found to form a complex in vitro with four other proteins Nap1 (Nck-associated protein, Kette in Drosphila), PIR121 (p53 inducible mrna, CYFIP in Drosphila), Abi2 (Abl interactor, Abi in Drosphila) and HSPC300 each of which had previously been associated with cytoskeletal functions [30]. CYFIP is probably the link between the pentrameric complex 14
15 and Rac, as it bears homology to p140sra1, a Rac effector. CYFIP is also likely to mediate the indirect interaction between Nck and Kette, which itself directly interacts with Abi. Finally, Abi links CYFIP/Kette to SCAR by binding to its N-terminal SHD (SCAR homology domain). HSPC300 also interacts with Abi and the SHD of SCAR. The incorporation of the five proteins into the complex has been proposed to maintain its stability. In Drosophila cells, RNAi-mediated ablation of any member of the complex, except HSPC300, correlates with the degradation of SCAR via the proteasome pathway [31]. The sequence of events that leads to activation of SCAR in not yet clear, but two models exist and are described in Scheme 4. According to one scheme, the binding of Rac to CYFIP or the binding of Nck to Kette causes the complex to break down to two sub-complexes. The suppressing CYGIP/Kette/Abi complex splits from the active SCAR/HSPC300 sub-complex, which promotes actin nucleation, but is then rapidly degraded. The other scheme suggests that the complex does not split, but the binding of the activator changes the conformation of SCAR enabling it to become active where Rac or Nck are localized [32]. Several studies link the fusion process to changes that occur in the actin cytoskeleton during the association between founder and FCM cells. Mutants in kette, which encodes a member of the SCAR/WAVE complex of proteins that is known to control actin polymerization by the activation of the Arp2/3 complex, were shown to exhibit myoblast fusion arrest [20], as well as some mutations in the Arp2/3 complex itself [33]. EM analysis revealed that kette-mutant myoblasts are arrested at the electron dense plaque stage, and fail to form membranal pores. Next, four recently published papers [33-36] deal with the role of actin polymerization, promoted by another Arp2/3 activator protein, WASp, in myoblast fusion. Wsp mutants were also shown to exhibit a fusion arrest, but the stage at which Wsp functions is a matter of debate. Data from our lab suggested that in Wsp mutants small pores open in the membrane between pairs of fusing cells, but pore extension and membrane breakdown are not achieved. It also seems that none of the common activators of WASp participate in the process, and it is the localization of WASp by its adaptor protein, Drosophila-WASp Interacting Protein (D- Wip), at the fusion site, that enables fusion to continue normally[35]. D-wip was shown 15
16 to interact with the cytoplasmatic portion of the fusion receptors, Duf and SNS, which likely accounts for the recruitment of WASp to the site of fusion. The rich biochemical data gathered about these proteins makes it appealing to investigate their function in a biological context. The fusion of myoblasts is a developmental process utilizing both the WASp and SCAR pathways, thus it might enable us to translate the knowledge from cell culture to the whole organism. The initial phase of my thesis research was dedicated to study the interplay between these two actin nucleators in vivo during myoblast fusion (Chapter 2.) Metamorphosis and the Formation of the Adult Musculature Adult somatic muscles are formed by the fusion of cells that originate from APs, myoblasts that are set-aside in the embryo and proliferate during larval life. At the onset of metamorphosis most larval muscles begin to histolyse. At this time, adult myoblasts migrate to specific ectodermal locations. There they fuse to form syncitial muscle fibers 3. The architecture of the adult somatic muscles is different from that of the larval somatic muscles. In the larva, each somatic muscle is a single multinucleated fiber whereas in the adult fly, similar to the situation in many skeletal muscles of vertebrates, each muscle is a collection of fibers. While the adult fly body contains many somatic muscles, this work focuses on the dorsal longitudinal flight muscles (DLMs), thoracic muscles that are positioned in the thorax and occupy most of it. Their contractions are the key energy-generating force that allows the insect to fly. In the embryonic thorax APs are present as a cluster of 6-7 cells in each hemisegment, and remain tightly associated with the imaginal disc primordia (groups of cells set aside in the embryonic and larval life, which give rise to adult epidermal structures like wings or legs during metamorphosis) [1]. From the beginning of the second larval instar, these adult precursor cells proliferate, and associate with the imaginal discs and nerves. With the onset of metamorphosis, the wing discs evert to release the myoblasts, which then migrate to specific epidermal positions and fuse, to form the thoracic muscles. In general, myoblasts associated with each imaginal disc appear to contribute to muscles that are associated with the epidermal structure formed by that disc [37]. 16

17 Major muscle groups of the thorax are the DLMs, and the dorso-ventral flight muscles (DVMs). The development of these two muscle groups is fundamentally different from each other. During metamorphosis three dorsal muscles of the larva in each mesothoracic hemisegment are spared from histolysis. These three fibers function as templates for the future DLMs to which the incoming myoblasts fuse. Each template subsequently splits into two, to generate the six fibers that constitute the DLMs (Scheme 4). On the other hand, the DVMs are formed de novo as a result of fusion of imaginal myoblasts with founder cells that have also migrated from the imaginal discs[38-40]. Scheme 4. Schematic representation of the development of the indirect flight muscles (IFMs) and the abdominal muscles. The IFMs are anatomically and functionally subdivided into two distinct groups: the dorsal longitudinal muscles (DLMs) and the dorsoventral muscles (DVMs). (A) The precursor myoblasts (yellow) for the IFMs are attached to the wing disc during the larval life. At the onset of metamorphosis larval muscles begin to histolyse, with the exception of three oblique muscles. (B) By 12 hours after pupa formation (APF), histolysis is complete and the three larval templates are clearly visible. By this stage, myoblasts migrate into the muscle-forming regions. (C) The larval templates start splitting by 13 hours APF and the process is complete by hours APF. At the same time, the DVMs form by the de novo fusion of myoblasts. (D) By 24 hours APF, the DLMs and the DVMs are complete. Muscle formation in the abdomen occurs later than in the thorax. The abdominal muscles develop from an adult myoblast pool (yellow) associated with the segmental and intersegmental nerves (A). (A-D) During early pupal stages (from 0-24 hours APF), myoblasts proliferate and migrate out along the nerves. (E) Fusion of these myoblasts begins by hours APF. (F) The arrangement of the DLMs, the DVMs (I, II and III) and the dorsal muscles of the first abdominal segment in an adult fly: by 50 hours APF, the pattern of the adult muscles is largely complete. Adapted from [41]. 17
18 1.6. Notch signaling pathway It is clear that cell communication has an essential role in the process of cell fusion. Cell communication triggers many differentiation processes and is crucial in the cascade of events that lead to the adoption of a fusogenic cell fate. One of the signaling pathways that participates at numerous steps during the differentiation into myoblasts is the Notch pathway. The Notch signaling pathway is a highly conserved cell signaling system present in most multicellular organisms [42]. Drosphila has a single notch receptor (N) and mammals possess four different Notch receptors, referred to as NOTCH 1-4. The Notch receptor is a single-pass transmembrane protein. It is a hetero-oligomer composed of a large extracellular portion, which associates in a non-covalent interaction with a smaller portion of the Notch protein composed of a short extracellular domain, a single transmembrane-pass, and a small intracellular region. Ligands binding to the extracellular domain induce proteolytic cleavage and release of the intracellular domain, which then enters the nucleus to trigger gene expression [43]. The Notch receptor is normally triggered only by direct cell-to-cell interaction, since the Notch ligands are also transmembrane proteins. This is a unique feature of the pathway, allowing groups of cells to self-organize themselves. This is termed lateral inhibition, and plays a role in many Notch signaling events. Lateral inhibition is one of the major themes of development. The process of lateral inhibition and cell selection is repeated hundreds of times in Drosophila, with differentiation that takes place in nearly every kind of tissue. For example: in the differentiation of FCMs and FCs, limiting the number of neural precursors, limiting the growth of Malpighian tubules, and regulation of the growth of the ovary. Lateral inhibition is a process whereby a single cell is fated to differentiate through the interaction of Notch-Delta, while neighboring cells simultaneously retain their undifferentiated state. A state of competition is imposed upon a cluster of cells. A single cell, seemingly selected at random, is the one with the highest density of ligand, caused by stochastic fluctuation in expression of the ligand. The selected cell signals to its adjacent cells, preventing them from down-regulating the Notch receptor, however terminating their ligand expression. Example for such a regulatory loop between Notch 18
19 and its ligand Delta can be found in studies concerning the physical spacing of precursors for the microchaetes of the peripheral nervous system. Activation of Notch leads to repression of the achaete-scute genes, which are themselves known to regulate transcription of Delta; this regulation may perhaps be direct [44]. Maturation of the Notch receptor involves cleavage at the prospective extracellular side during intracellular trafficking in the Golgi complex [45]. This result in a split protein, composed of a large extracellular domain linked by disulphide bonds to the smaller transmembrane and intracellular domains. Binding of ligand promotes two proteolytic processing events. As a result of proteolysis, the intracellular domain is liberated and can enter the nucleus to engage other DNA-binding proteins and regulate gene expression. The Notch ligands are also single-pass transmembrane proteins and are members of the DSL (Delta/Serrate/LAG-2) family. In Drosophila, there are two ligands named Delta and Serrate. In mammals, the corresponding names are Delta-like and Jagged. In mammals there are multiple Delta-like and Jagged ligands, as well as possibly a variety of other ligands, such as F3/contactin [46]. The Notch extracellular domain is composed primarily of small cysteine knot motifs called EGF-like repeats [47]. Notch 1, for example, has 36 of these repeats. Each EGF-like repeat is composed of approximately 40 amino acids, and its structure is defined largely by six conserved cysteine residues that form three conserved disulfide bonds. Each EGF-like repeat can be modified by O-linked glycans such as O-glucose or an O-fucose at specific sites [48]. Addition of these sugars is absolutely necessary for Notch function, and, without the enzyme to add O-fucose, all Notch proteins fail to function properly. As yet, the manner by which the glycosylation of Notch affects function is not completely understood. The O-glucose on Notch can be further elongated to a trisaccharide with the addition of other sugars. This is mediated by GlcNAc-transferases. In Drosophila one such enzyme has been identified, Fringe [49]. Following full glycosylation, Notch signals strongly when it interacts with the Delta ligand, but has markedly inhibited signaling when interacting with the Serrate/Jagged ligand [50]. The means by which this addition 19
20 of sugar inhibits signaling through one ligand, and potentiates signaling through another, is not clearly understood. Once the Notch extracellular domain interacts with a ligand, an ADAMfamily metalloprotease cleaves the Notch protein just outside the membrane [51]. After this first cleavage, an enzyme called γ-secretase cleaves the remaining part of the Notch protein just inside the inner leaflet of the cell membrane of the Notch-expressing cell. This releases the intracellular domain of the Notch protein, which then moves to the nucleus, where it can regulate gene expression by activating the transcription factor CSL (Suppressor of Hairless, in Drosophila). The interaction of the intracellular portion of Notch with the CSL factors, reduces their affinity to transcriptional repressor such as hairless [52], and transforms them to transcriptional activators of Notch-target genes Cis-inhibition of Notch signaling Notch receptors in a given cell are activated by cell surface ligands in neighbouring cells, but can also be inhibited by the ligands present within the same cell. This process is known as cis-inhibition of Notch. Additionally, reciprocal cis-inhibition of the ligands by Notch has also been observed (reviewed in [53]). Early genetic experiments in Drosophila uncovered surprising relationships between Notch and its ligand Dl. In certain contexts, double heterozygocity for ligand and receptor suppressed the individual phenotypes of each other while gene duplication of the ligand enhanced receptor loss of function phenotypes and vice versa [54-56]. These interactions suggested that the ratio between the ligand and the receptor concentrations can influence the signaling outcome. In 1997, two papers [57, 58] demonstrated, for the first time, that ligands actually display a cis-inhibitory effect on Notch, in order to properly specify the wing margin. The molecular mechanism underlying cis-inhibition of Notch by its ligands is only now beginning to emerge. Several lines of evidence indicate that it involves direct ligand receptor interaction. In vivo structure function studies in Drosophila showed that the receptor-binding domain of Ser is required for cis-inhibition [59]. A recent structural analysis further suggested that Notch may interact with the DSL domain of its ligands via 20
21 two modes of interaction, referred to as parallel and antiparallel binding: interaction of Ser with Notch in trans would follow antiparallel binding, leading to activation of Notch, whereas interaction in cis would be mediated by parallel binding, resulting in inhibition of Notch [60]. Interaction of Notch with its ligands in cis may inhibit signalling by forming complexes that are inactive for signal reception, thereby titrating Notch. 21
22 2. The SCAR and WASp nucleation promoting factors act sequentially to mediate Drosophila myoblast fusion Boaz Gildor, R ada Massarwa, Ben-Zion Shilo and Eyal D. Schejter Dept. Molecular Genetics, Weizmann Institute of Science, Rehovot 76100, ISRAEL 2.1. Abstract The actin nucleation-promoting factors SCAR/WAVE and WASp, together with associated elements, mediate formation of muscle fibers via myoblast fusion during Drosophila embryogenesis. Our phenotypic analysis, following disruption of these two pathways, suggests that they function in sequential fashion. SCAR activity is required prior to the formation of pores in the membranes of fusing cells, while WASp promotes expansion of nascent pores and completion of the fusion process. Genetic epistasis experiments are consistent with this step-wise temporal progression. Our observations further imply a separate, Rac-dependent role for the SCAR complex in promoting myoblast migration. In keeping with sequential utilization of the two systems, we observe abnormal accumulations of filamentous actin at fusion sites when both pathways are disrupted, resembling those present when only SCAR complex function is impaired. This observation further suggests that actin filament accumulation at fusion sites may not depend on Arp2/3 activity altogether. 22
23 2.2. Introduction The evolutionarily-conserved Arp2/3 complex acts as a key mediator of actin polymerization in living cells, and the forces generated by Arp2/3-based branched polymerization provide a mechanistic basis for a diverse array of fundamental cellular processes [61, 62]. Nucleation of actin polymerization by the Arp2/3 complex is commonly stimulated by nucleation promoting factors (NPFs), which serve as critical links between signal-transduction pathways and remodeling of the actin-based cytoskeleton. Members of the WASp and SCAR/WAVE protein families are considered to be the two major NPFs mediating Arp2/3 function [63]. While the molecular basis for Arp2/3 activation is shared by both NPFs, these modular proteins are subject to distinct modes of regulation by components of signaling pathways. Thus, while Arp2/3-based actin polymerization constitutes a common outcome, the two NPFs are often utilized in distinct settings [64]. The fruitfly Drosophila melanogaster provides a system for assessing the physiological roles of the Arp2/3 machinery and its associated NPFs, as it possesses single WASp (Wsp; [65]) and SCAR/WAVE (SCAR;[66]) homologs. While genetic analyses have commonly identified distinct, non overlapping roles for SCAR and Wsp during development, a series of recent studies has revealed requirements for both NPFs in the process of embryonic myoblast fusion. Formation of functional muscle fibers in Drosophila embryos is based on successive rounds of fusion between founder-cell myoblasts (FCs) and the more numerous class of fusion-competent myoblasts (FCMs). Recognition and attachment between FCs and FCMs, mediated by cell-type-specific adhesion molecules, sets in motion a series of events that culminates in opening of fusion pores and eventual breakdown of the aligned myoblast membranes. A role for SCAR in this process was suggested both by study of Kette (NAP1), a conserved member of the SCAR regulatory complex [20], as well as by direct assessment of SCAR function [67, 68]. Complementing studies of Wsp, and of the Wsp regulatory element D-WIP/Sltr/Vrp-1 [34, 35, 69], have put forward strong evidence that both types of Arp2/3 NPFs provide essential contributions to myoblast fusion in Drosophila embryos. 23
24 The shared involvement of both SCAR and Wsp in myoblast fusion, allowed us to assess the functional contributions of Arp2/3 NPFs within a common physiological framework. We find that SCAR is required for myoblast migration as well as myoblast fusion, in keeping with the pleiotropic nature of its function throughout Drosophila development [66]. Furthermore, we demonstrate that SCAR and Wsp act in a sequential manner to promote myoblast fusion. Our findings therefore imply that the two NPF systems are responsible for distinct mechanistic functions, even when operating in close spatial and temporal correspondence. 24
25 2.3. Results and Discussion Multiple roles for the SCAR complex during embryonic myogenesis To assess and compare the defects in embryonic myogenesis resulting from disruption of SCAR and Wsp function, we examined embryos bearing strong loss of function alleles of kette and D-WIP, respectively (Fig 1). Genetic disruption of these regulatory elements provides the most severe myogenic phenotypes associated with zygotic Arp2/3 NPF loss-of-function (see Supplementary information for further discussion and for detailed analysis of SCAR-complex phenotypes). As previously reported, both D-WIP (Fig 1B) and kette (Fig 1C) stage 16 mutant embryos display hallmark abnormalities in muscle development associated with defects in the myoblast fusion process, including strikingly thin muscle fibers and numerous unfused myoblasts, which are nearly absent from wild-type embryos at this stage. Closer examination reveals clear phenotypic distinctions between the D-WIP and kette mutant embryos (Fig 1D, E, G, H). These are most readily apparent in both the distribution and morphology of the unfused myoblasts. In D-WIP embryos, unfused myoblasts are typically clustered and attached to myotubes, and approximately half of those that remain unattached display a migratory, tear-shaped morphology, in which a broad lamellipodium is extended in the direction of cell movement. In kette mutants, however, unattached myoblasts are much more abundant, and commonly retain a rounded morphology, consistent with the involvement of SCAR/WAVE elements in regulating morphological transitions and associated changes in migratory behavior [70]. In order to investigate this issue further, we used anti-scar antibodies to localize the endogenous SCAR protein in the myogenic mesoderm (Fig 2). We performed this analysis in wild-type (Fig 2A-A ) as well as D-WIP mutant embryos (Fig 2B-B ), in which the fusion arrest enriches for myoblasts that are at the various stages of migration and fusion. SCAR protein is found to be expressed in both myotubes and individual FCMs, where it displays a dynamic pattern of localization. SCAR is uniformly distributed within the cytoplasm of round, pre-migratory myoblasts. However, as myoblasts acquire a tear-shaped morphology, SCAR becomes asymmetrically distributed, and is conspicuously enriched in the lamellipodial extension of nearly all tear-shaped cells. SCAR remains concentrated within this structure as the migrating myoblasts orient and 25
26 move towards the myotubes. Myotube-myoblast attachment leads to establishment of fusion sites, which can be visualized by accumulation of the myotube-specific fusion receptor Duf (Fig 2A, 2B ). SCAR is enriched at these sites. The localization pattern of SCAR protein during these phases of myogenesis is therefore consistent with functional roles in both migration and fusion. Signaling via the small GTPase Rac provides a primary pathway for stimulation of SCAR/WAVE complexes and their associated NPF activity. We therefore examined embryos bearing strong loss-of-function alleles in all three Drosophila rac orthologs, rac1, rac2 and mtl (rac triple mutants). As previously reported [68, 71], compromising rac function in this manner results in strong impairment of myoblast fusion (Fig 1F). Furthermore, the abundance and rounded morphology of unfused myoblasts are reminiscent of the defects associated with disruption of SCAR complex activity (Fig 1E). Further examination of rac-triple mutant embryos revealed significant abnormalities in the pattern of SCAR protein localization during myoblast fusion and migration (Fig 2C-C ). SCAR becomes uniformly distributed in myoblasts and myotubes, and fails to exhibit any specific pattern of sub-cellular localization. This holds true both for the asymmetric distribution within unattached myoblasts, regardless of their morphology, as well as the enrichment at fusion sites. rac function thus appears to be essential for proper localization of the SCAR protein in both migrating and fusing myoblasts. 26
 Full dorsal views of stage 16 wild-type (A), D-WIP D30 (B), and kette J4-48 (C) embryos stained with anti- MHC (green), to reveal the embryonic musculature.")
27 Figure 1. Distinct myogenic phenotypes result from disruption of the Wsp and SCAR pathways. (A- C) Full dorsal views of stage 16 wild-type (A), D-WIP D30 (B), and kette J4-48 (C) embryos stained with anti- MHC (green), to reveal the embryonic musculature. (D-F) Higher magnification images of D-WIP D30 (D), kette J4-48 (E) and rac1 rac2 mtl triple-mutant (F) embryos, stained with anti-mhc (green), as well as for the rp298/duf-lacz reporter [72], expressed in myotube nuclei (red). While the muscle pattern of both NPF pathway mutants is severely disrupted, unfused myoblasts in the D-WIP embryos (B,D) are mostly clustered and attached to small myotubes, while those in the kette (C,E) and rac (F) embryos are far more dispersed. Arrows in panels D-F point to representative unattached myoblasts, which are commonly tear shaped in the D-WIP embryo, but retain a rounded, nonmigratory morphology in the kette and rac embryos. Scale bars (in A,D)= 50 μm. (GH) Quantitative assessment of myogenic phenotypes. The proportion of unattached myoblasts (G) and of those displaying a migratory morphology (H) was determined in the different mutant backgrounds. n represents the total number of myoblasts counted in at least three different embryos of each genotype. 27
 A stage 13 wildtype twist-gal4::uas-gfpmyr embryo stained with anti-gfp (A, green) to visualize the contours of myoblasts, anti-scar (A, red) and anti-duf (A, blue), which marks myotube")
28 Figure 2. Localization pattern of SCAR in migrating and fusing myoblasts. (A-A ) A stage 13 wildtype twist-gal4::uas-gfpmyr embryo stained with anti-gfp (A, green) to visualize the contours of myoblasts, anti-scar (A, red) and anti-duf (A, blue), which marks myotube surfaces. The asterisk marks a rounded (pre-migratory) myoblast in which SCAR is uniformly localized in the membrane, an arrow points to a tear shaped, migrating myoblast, where SCAR localizes to the lamellipodium, and the arrowhead to a myoblast/myotube fusion interface. (B-B ) A stage 14 D-WIP D30 embryo. Anti-MHC (B, green) was used as the general myoblast marker. SCAR localization within the enriched population of unfused myoblasts resembles that seen in wild-type. Cells are marked as in A, A. (C-C ) A rac triple mutant embryo stained as in (B-B ). SCAR is uniformly distributed within the unfused myoblasts, all of which display a rounded morphology. Scale bar= 10 μm. Distinct, sequential roles for SCAR and Wsp during myoblast fusion Phenotypic distinctions among Drosophila embryonic myoblast fusion mutants are readily apparent, following examination by electron microscopy (EM), which has identified fusion-specific cellular structures, and has enabled categorization of the fusion process into discrete steps [9]. We thus chose to pursue EM-level analysis of SCAR complex and D-WIP/Wsp mutants, to assess the consequences of separate and simultaneous genetic disruption of Arp2/3 NPF activity. 28
29 We used embryos homozygous for the amorphic kette J4-48 allele [20, 73] to represent strong SCAR complex loss-of-function conditions, and Wsp 3D3 homozygotes to represent disruption of Wsp function. Wsp 3D3 encodes an inactive Wsp gene product, that also interferes with the activity of maternally-contributed Wsp ([69]; see Supp. Information for additional phenotypic analysis). kette J4-48 embryos (Fig 3A) display all of the established cytological features of fusing myoblasts [9], including the electron dense plaques that appear in the aligned membranes of attached cells just prior to opening of fusion pores (Fig 3A, inset). Importantly, however, in none of ~100 attached myoblast pairs examined by EM, were any discontinuities indicative of fusion pore formation observed in the paired myoblast membranes of kette mutants. These observations imply, therefore, that SCAR complex stimulation of Arp2/3-based actin polymerization is required at a relatively advanced stage of the fusion process, perhaps as an accessory to the establishment of fusion pores. A clearly distinct picture is revealed following examination of the arrested myoblast fusion in Wsp 3D3 homozygous embryos (Fig 3B). Roughly half of all attached myoblast pairs in these embryos (n~100) display multiple, small ( nm) breaches distributed along the length of apposed myoblast membranes. This is fully consistent with our previous report on embryos lacking either Wsp or D-WIP function [35], and implies a requirement for the Wsp-associated molecular machinery during expansion of nascent fusion pores. These phenotypic differences therefore allow for assessment of an epistatic relationship between the SCAR and Wsp systems. Towards this end we examined homozygous kette J4-48 Wsp 3D3 double-mutant embryos [69], where both NPF systems are compromised simultaneously. We find that the double-mutant phenotype is identical to that of kette J4-48 alone: fusion-related structures are present, but none of the apposed membranes (in ~50 myoblast pairs examined) show any sign of fusion pore formation (Fig 3C). The kette phenotype is therefore epistatic to Wsp, supporting a step-wise scenario, in which SCAR complex activity precedes the requirement for D-WIP/Wsp. We have previously demonstrated that the presence of partial membrane openings between attached myoblasts, such as in D-WIP mutant embryos, can be verified by monitoring transfer of cytoplasmic material from one cell to the other. We have now used this assay to complement the EM observations on kette and Wsp mutant embryos. While 29
30 no indication of GFP transfer from myotubes to myoblasts can be detected in kette J4-48 mutants (Fig 3 D-D ), at least half of the attached myoblasts in Wsp 3D3 homozygotes clearly contain GFP (Fig 3E-E ), consistent with the achievement of cytoplasmic continuity. Strikingly, and in full correspondence with the EM observations, we fail to observe any instance of GFP transfer in homozygous kette J4-48 Wsp 3D3 double-mutant embryos (Fig 4F-F ). This independent demonstration of genetic epistasis underscores the sequential requirement for the two Arp2/3 NPF systems during embryonic myoblast fusion. We wish to stress the significance of these observations, in light of the recent controversy surrounding the phenotype of embryos defective in D-WIP function [74]. Our data continues to demonstrate formation of small membrane pores, and a corresponding transfer of cytoplasmic GFP between attached myoblasts, specifically in D-WIP and Wsp mutant embryos, but not in any other mutant background. Furthermore, we now demonstrate the suppression of both the perforated membrane phenotype and of GFP transfer in kette J4-48 Wsp 3D3 double mutants. We believe that these observations provide significant support for our assertion that when D-WIP/Wsp function is compromised, fusion is arrested only after cytoplasmic continuity between fusing myoblasts has been achieved. 30
 Closely aligned and fully intact membranes in a kette J4-48 mutant embryo. Inset shows an electron dense plaque (arrowhead), commonly observed in the apposed myoblast membranes in these embryos.")
31 Figure 3. SCAR and Wsp pathway mutants arrest at distinct stages of the myoblast fusion process. (A-C) EM micrographs of attached pairs of myoblasts. (A) Closely aligned and fully intact membranes in a kette J4-48 mutant embryo. Inset shows an electron dense plaque (arrowhead), commonly observed in the apposed myoblast membranes in these embryos. (B) Multiple gaps (arrows) form in the apposed myoblast membranes of a Wsp 3D3 mutant embryo. (C) Attached myoblast membranes remain intact in a kette J4-48 Wsp 3D3 double-mutant embryo. Fusion-related structures, including electron-dense plaques (inset), are present, as in the kette mutant alone. (D-F) Monitoring cytoplasmic transfer in SCAR and Wsp pathway mutants, following expression of GFP in myotubes via rp298/duf-gal4::uas-gfp (Menon & Chia, 2001). Mutants shown include kette J4-48 (D- D ), Wsp 3D3 (E-E ), and the kette J4-48 Wsp 3D3 double (F-F ). All genotypes were stained with anti-mhc (grey), to determine myoblast cell outlines, anti-duf (red), to mark sites of arrested fusion, and anti-gfp (green). Asterisks (E ) mark GFP-positive FCMs in Wsp 3D3. Scale bars=200 nm (A),10 μm (D). 31
32 32
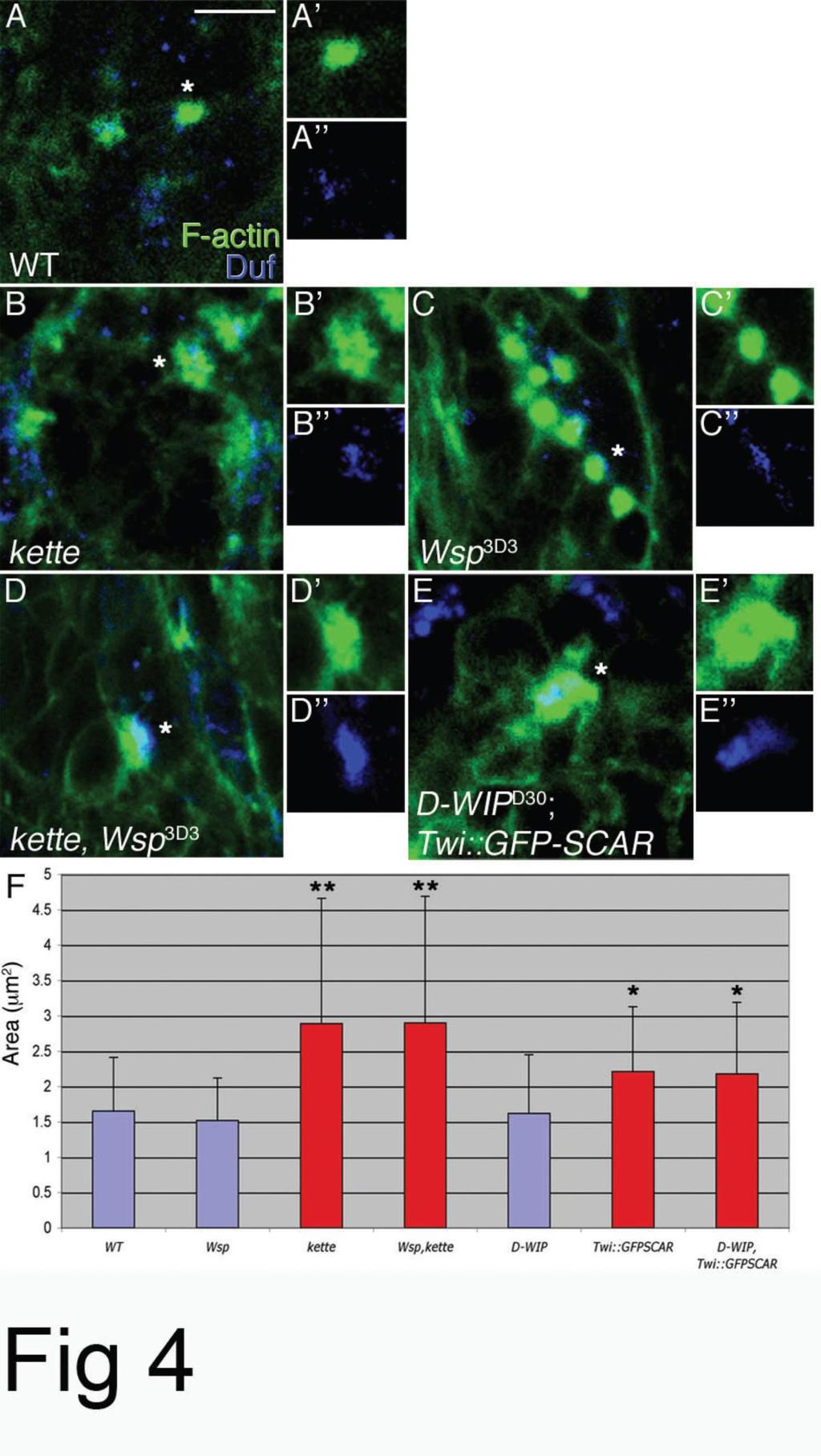
33 Figure 4. F-actin foci form and persist despite disruption of both the SCAR and Wsp NPF pathways. (A-E) Phalloidin-staining structures (green) in stage 14 wild-type (A), kette J4-48 (B), Wsp 3D3 (C), kette J4-48 Wsp 3D3 (D) and D-WIP D30 twist-gal4::gfp SCAR (E) embryos. All embryos were also stained with anti- Duf (blue), to visualize myotube surfaces and fusion sites, and anti-mhc, to visualize myoblasts (not shown). For each genotype, isolated phalloidin and anti-duf stainings of single representative F-actin foci (asterisks) are shown as well. Note enlarged and irregular morphology of foci in kette and double-mutant embryos, as opposed to the round shape of wild-type and Wsp foci. Scale bar= 10 μm. (F) Bar diagram showing quantification of F-actin foci diameters. Asterisks mark mutant genotypes where the difference from wild-type was statistically significant. Blue and red coloring are used to, respectively, designate round vs. spread-out, irregular focus morphologies. The formation of actin foci at fusion sites is not mediated by SCAR and Wsp NPF activity Several recent studies have documented conspicuous F-actin concentrations, commonly referred to as actin foci, at the interface of fusing myoblasts in Drosophila embryos [34, 68, 75]. The temporal profile of appearance and dissolution of the actin foci in wild-type embryos closely parallels the dynamics of myoblast fusion [68], suggesting that these structures play an important role in the process. The substantial functional requirements for components of the Arp2/3 machinery in embryonic myoblast fusion, naturally raise the possibility that construction of the fusion associated foci depends on Arp2/3-based actin polymerization. We examined this issue using single and double-mutant allelic combinations of zygotic loss-of-function situations for both NPF systems (Fig 4). Consistent with published reports [68], we find that disruption of SCAR-complex function results in formation of enlarged, irregularly shaped foci at sites of myotube myoblast attachment. Homozygous kette J4-48 mutant embryos display foci whose area is nearly twice the size of wild-type foci ( μm2 vs μm2, P= 5.66E-08, n=67). Enlarged foci ( μm2, P=0.007, n=162) are also characteristic of embryos expressing the dominant-negative GFP-SCAR construct in muscle tissue. We chose to disrupt SCAR function in this manner, since it generates stronger fusion phenotypes than those obtained with zygotic SCAR alleles (Supp. Fig 1). To assess the requirement for Wsp activity, we examined actin focus morphology in Wsp 3D3 homozygotes, and in embryos homozygous for D-WIP D30 [35], a complete deletion of the D-WIP locus. We find that actin foci form under both these circumstances, but do not grow to larger dimensions than those present in wild-type embryos ( μm2 and μm2, respectively; see also [76]). 33
34 We next sought to determine whether actin foci form following simultaneous disruption of both NPF systems. Towards this end we examined both kette J4-48 Wsp 3D3 and twist-gal4::uas-gfp-scar; D-WIP D30 double-mutant embryos. In both cases we observed enlarged actin foci ( μm2, P=3.72E-07 and μm2, P=0.015, respectively), resembling those present in the corresponding single mutants of SCARcomplex elements. The molecular basis for construction of fusion associated actin foci remains, therefore, unknown. Furthermore, the analysis of single and double-mutant combinations is fully consistent with the epistatic relationship between the NPF systems derived from the EM and GFP transfer studies, supporting our assertion that the SCAR and D-WIP/Wsp systems act separately and sequentially during myoblast fusion. In the current study, we have assessed and compared the contributions of the two primary Arp2/3 NPFs, SCAR/WAVE and WASp, to the process of myoblast fusion during Drosophila embryogenesis. Our analysis suggests a scenario wherein the SCAR complex is utilized twice during embryonic myogenesis, initially promoting migration of myoblasts towards growing myotubes in a Rac-dependent manner, and later during the fusion process itself. The Wsp NPF system, on the other hand, is assigned a single, independent role, subsequent to fusion-pore formation. A model summarizing our interpretation of the myogenic functions involving Arp2/3 NPFs is shown in Fig 5. A central feature of this scenario, which we believe to bear significance for the in-vivo roles ascribed to the Arp2/3 system, is that NPFs that use a common molecular mechanism to target the same actin-polymerization machinery, and do so in close spatial and temporal proximity, can still mediate distinct cellular functions. Deciphering the activation mechanisms that allow SCAR and Wsp to carry out distinct roles in muscle fusion via the common Arp2/3 actin nucleator, is a future challenge. 34

35 Figure 5. Model of the functional contributions of the SCAR and WASp pathways to myoblast fusion in Drosophila embryos. A SCAR-complex dependent morphological transition underlies acquisition of migratory behavior by myoblasts, which are now capable of attaching to founder cells/myotubes. Upon initiation of fusion, the SCAR complex is again utilized to promote formation of pores in the apposed cell membranes. Once initial pores are formed, Wsp, recruited by D-WIP, mediates pore expansion, presumably via forces derived from Arp2/3-based actin polymerization. Note: An alternative model for the contribution of the two pathways has been recently proposed [67]. 35
36 2.4. Methods Drosophila genetics See specific references and Flybase [available at for details concerning all genetic loci and mutant alleles described throughout this study. Embryo processing, immunohistochemistry and microscopy Embryos were processed for viewing by fluorescent and electron microscopy as previously described [35]. See Supp. Info. for additional technical details and list of antibodies used. Fluorescent images were collected on a Zeiss LSM510 confocal system. Actin foci measurements were performed using the Overlay function of the Zeiss LSM software (see also [68]). 36
37 2.5. Acknowledgements We are grateful to M. Baylies, P. Fischer, M. Frasch, C. Klämbt, D. Menon, S. Önel, R. Renkawitz-Pohl, M. Ruiz-Gomez, T. Volk and the Bloomington Stock Center for stocks and reagents. We thank Michal Haskel for contributing to SCAR-complex phenotypic analysis, Orit Bachar for drawing the model in Figure 5, and all members of our lab for discussion and support. This study was financially supported by grants from the Mary L. Ralph Designated Funds and the Israel Science Foundation to B-Z. Shilo and E. Schejter. B-Z. Shilo is an incumbent of the Hilda and Cecil Lewis chair in Molecular Genetics. 37
38 2.6. Supplemental Information to: " The SCAR and WASp nucleation promoting factors act sequentially to mediate Drosophila myoblast fusion" I. SCAR system 1. Assessment of phenotypic consequences resulting from zygotic disruption of SCAR-complex function. Zygotic disruption of the D-WIP locus provides a good means to assess Wsp pathway function during myoblast fusion, given the exclusively zygotic mode of D-WIP gene expression, and the close similarities between D-WIP and Wsp myogenic phenotypes [35]. Use of the Wsp 3D3 allele (see below), provides an alternative method to achieve this end. However, choosing an appropriate genetic background for parallel study of SCAR NPF function is less straightforward, given apparent variabilities in maternal contributions and lack of complete information regarding SCAR regulatory factors. We therefore surveyed phenotypes resulting from complete zygotic disruption of all Drosophila homologs of SCAR regulatory complex elements, as well as SCAR itself. Nuclei of the dorsal embryonic muscle DAI express the transcription factor Even-skipped (Eve), and determination of the number of such Eve expressing nuclei serves as an established and sensitive assay for the severity of fusion defects [77]. As shown in Supp. Table 1, zygotic disruption of kette, the Drosophila NAP-1 homolog, provides the strongest phenotype (see also [20]), and this background was therefore chosen for the analyses presented in the main text. 2. Generation of a SCAR dominant-negative allele. The pleiotropic nature of SCAR complex phenotypes motivated us to obtain specific functional impairment of SCAR during myogenesis, based on a dominant negative approach and the GAL4-UAS system for regulated expression [78]. An initial attempt involved expression of a SCAR protein construct lacking the C-terminal VCA Arp2/3 and actin binding domains. This approach, which proved successful with Wsp [35], failed in this case (data not shown), possibly due to instability of SCAR when subjected to structural alterations. However, a second construct, in which the full SCAR coding region is fused to GFP at its N terminus, was found to cause a severe fusion arrest in the majority of embryos (Supp Fig 1), when expressed in developing muscles using the muscle specific driver Mef2-GAL4 [79]. The fusion arrest phenotype can be overcome by co-expression of the intact SCAR protein 38
39 (Supp Fig 1M). We therefore suggest that the GFP-SCAR construct generates a stable, inactive form of the SCAR NPF, which competes with the endogenous SCAR protein in binding to the other SCAR-complex members. Supp Table 1. Quantification of myoblast fusion arrest in SCAR-complex mutants. The DA1/Eve assay was applied to wild-type embryos and embryos homozygous for the following genotypes: kette J4-48, an amorphic kette allele [20, 73](kette is the Drosophila homolog of vertebrate NAP-1); SCAR 37, a full deletion of the SCAR locus [66]; Df(3R)red3l, a chromosomal deficiency spanning the abi locus at cytogenetic band 88A9; Df(2R)Chi g230, a chromosomal deficiency spanning the SIP1/HSPC300 locus at cytogenetic band 60B4; and Sra , a null allele of the Sra-1/CYFIP gene [80]. The kette mutant displays a severe fusion phenotype (see also Supp. Fig 1 below), while the phenotypes of the other mutants are substantially weaker. Asterisks mark values that differ in a statistically significant manner from wild-type. SD= standard deviation. 39
 document the dispersed, unfused myoblasts in four-segment wide embryonic sections of these genotypes.")
40 Supp Fig 1. Dominant-negative effects of GFP-SCAR. (A-L) Wild-type (A-C), kette J4-48 (D-F), SCAR 37 (G-I) and Mef2-GAL4::UAS-GFP-SCAR (J-M) stage 16 embryos. Panels A,D,G,J (anti-mhc, red) document the dispersed, unfused myoblasts in four-segment wide embryonic sections of these genotypes. Panels B,E,H,K (anti-eve, red) are representative images of the Eve/DA1 assay, quantified in Supp. Table 1. Images from an equivalent assay, quantifying the extent of fusion in muscle DO1 and using the Runt nuclear marker (blue), is shown in panels C,C,F,F,I,I,L,L. A bar diagram summarizing the DA1 and DO1 assays is shown in panel N. (M) Co-expression of UAS-SCAR overcomes the effects of UAS-GFP- SCAR (compare with panel J). II. Wsp system In the study, we made use of the Wsp 3D3 allele [69], to obtain strong Wsp loss-offunction circumstances without having to resort to germline clone analysis for elimination of the Wsp maternal contribution. To verify that the Wsp 3D3 allele can be used in this context, we ascertained that the dominant-negative activity of Wsp 3D3 activity is directed specifically towards Wsp, by demonstrating rescue of the Wsp 3D3 myogenic 40
. Supp Fig 2.")
41 phenotype by Wsp over-expression (Supp Fig 2). We then carried out Eve/DA1 and Runt/DO2 fusion-arrest assays on Wsp 3D3 homozygotes, as well as on kette J4-48 Wsp 3D3 double mutant embryos (Supp Fig 3). Supp Fig 2. Fusion-arrest phenotypes of Wsp 3D3. (A) MHC staining reveals strong fusion-arrest phenotype of a stage 15 Wsp 3D3 hemizygous embryo, which displays multiple unfused myoblasts clustered around short, thin myofibers. (B) Rescue of the Wsp 3D3 myogenic phenotype by expression of a full-length Wsp construct [65] in muscle tissue via twist-gal4. 41
 DO2 muscle nuclei expressing Runt (red) in Wsp 3D3 (C, C ) and kette J4-48 Wsp 3D3 (D, D ) mutants. DO2 Muscle contours can be seen via MHC staining (green) in panels C,D.")
42 Supp Fig 3. Quantification of the Wsp 3D3 fusion-arrest phenotype. (A,B) DA1 muscle nuclei expressing Eve (red) in Wsp 3D3 (A) and kette J4-48 Wsp 3D3 (B) mutants. (C,D) DO2 muscle nuclei expressing Runt (red) in Wsp 3D3 (C, C ) and kette J4-48 Wsp 3D3 (D, D ) mutants. DO2 Muscle contours can be seen via MHC staining (green) in panels C,D. A bar diagram summarizing these assays is shown in panel E. III. Methods- Immunofluorescence Primary antibodies and dilutions used in this study include: anti "-galactosidase (rabbit, 1:1,000); anti-duf (rat, 1:100); anti-gfp (mouse, 1:200; Roche); anti-mhc (rabbit, 1:500; gift from Paul Fisher, SUNY Stony Brook); anti-scar (guinea-pig, 1:100; Zallen et al., 2002). Secondary Cy2, Cy3 and Cy5-conjugated antibodies against the relevant species were from Jackson ImmunoResearch (West Grove, PA). Processing for experiments that included phalloidin staining involved manual devittelinization, 42
43 following fixation for 15 minutes on a heptane/formaldehyde:pbs (1:1) interface. Microfilaments were visualized by a 30 min incubation in 1 unit/ml Bodipy-conjugated phalloidin (Molecular Probes). Visualization of anti-scar required signal amplification as in Melen et al,
44 3. The Actin Nucleator WASp is Required for Myoblast Fusion During Adult Drosophila Myogenesis 3.1. Introduction The different developmental modes leading to formation of the adult Drosophila flight musculature (see chapter 1.5.) present opportunities to examine both fusion between individual myoblasts as well as between myoblasts and maturing fibers. While classic genetic approaches have proven to be a highly successful tool to study myoblast fusion during embryonic Drosophila myogenesis [24, 81], their application to the study of fusion during adult fly muscle development has been limited. This is due to both earlier functional requirements for many genes potentially involved in the process, and the syncytial nature of muscles, which restricts the usefulness of clonal analysis. In a recently published collaboration between the Shilo lab and the VijayRaghavan lab [82], a combination of genetic approaches to circumvent these difficulties, and identify essential contributors to adult myoblast fusion is described. Through use of classical mutants and gene knockdown by RNAi, a role for several critical NPFs in adult myoblast fusion is observed. Although Wsp performs essential roles throughout Drosophila development, flies lacking zygotic Wsp function survive until late pupal/early adult stages, since maternally- contributed Wsp gene products are sufficient for proper embryonic and larval development. Wsp mutant adult flies display severely deformed musculature. This deformity is caused due to a complete block in myoblast fusion in all different muscle types, independent of the developmental way in which they are generated. Both muscles in the DLM group which evolve from templates, and the de-novo generated abdominal muscles and DVMs, display a much reduced number of nuclei in them [82]. In agreement with rescue experiments done in the embryo [35], Wsp function can be rescued by expression of Wsp only on one of the fusion sides, either in the myoblasts or in the myotubes. Although Wsp is expressed ubiquitously in both cell types, an enrichment of the protein is observed on the tip of the fusing myoblast suggesting that Wsp normally localizes to the membrane regions actively involved in fusion. Furthermore, expression in myogenic cells of Wsp myr, a plasma membrane-tethered form 44
45 of Wsp [83], substantially rescues the Wsp mutant phenotype, underscoring the functional significance of targeting Wsp to muscle-cell membranes. D-WIP/Vrp1 is a WASp binding protein required for localization of the Wsp protein in embryonic myoblasts. The authors report that D-WIP is also required for adult myoblast fusion, however surprisingly D-WIP adopted a different function, as Wsp is normally localized in D-WIP mutants.. My contribution to this project was centered around the role of the SCAR complex in adult myoblast fusion. The primary phenotypic characterization of the SCAR complex RNAi was carried out by me, while the analysis of the formation of actin foci at the site of myoblast attachment was a collaboration in which I generated the recombinant flies and the staining and analysis were done by P.Mukherjee. These sections of the work are listed below in detail. 45
46 3.2. Results The Arp2/3 nucleation-promoting factor SCAR is required for adult myoblast fusion WASp-family proteins act as nucleation promoting factors (NPFs), stimulating the capacity of the conserved Arp2/3 protein complex to nucleate actin polymerization and generate branched microfilament arrays [61]. Metazoan cells commonly employ WASp-family proteins and the related SCAR/WAVE elements as the major Arp2/3 NPFs [84]. While the two classes of NPFs share a common mode of interaction with the Arp2/3 complex, they recognize and are activated by distinct molecular machineries [85]. Drosophila possesses single homologs for WASp (Wsp) and SCAR/WAVE (SCAR), which generally operate in separate developmental and cellular settings [66]. Embryonic myoblast fusion is exceptional in this regard, as both NPFs have been ascribed essential roles in the fusion process [20, 34, 35, 68, 69]. Direct comparisons, however, reveal separate and temporally distinct roles for the two Arp2/3 NPFs, where SCAR/WAVE acts prior to Wsp, raising questions regarding the mechanistic basis by which such functional distinctions are achieved [67, 86]. To determine the functional scenario for these factors during establishment of adult Drosophila muscles, we first sought to complement the above analysis of Wsp mutants, by disrupting the activity of SCAR and its associated molecular machinery in developing pupae. Unlike Wsp, the maternal contribution of SCAR and related elements is not sufficient to overcome zygotic requirements during embryonic and larval stages. We therefore chose to pursue restricted tissue and developmental-stage expression of UASbased RNA-interference (RNAi) transgenes [87, 88], as an alternative method of disrupting gene activity during establishment of the adult fly musculature. To verify the usefulness of this approach in the context of adult myoblast fusion, we first assessed the effects of expressing UAS-Wsp RNAi and UAS-D-WIP RNAi constructs, using the muscle specific mef2-gal4 driver. Severe myogenic phenotypes, resembling those obtained with loss-of-function alleles, were observed in both cases (Fig. 6A-C). 46
47 Incorporation of UAS-Dicer2, an established tool for enhancing RNAi activity [87], proved useful in this context, as it led to more severe and consistent defects in flight muscle formation. We therefore applied this protocol to examine the involvement of SCAR and associated elements in adult myogenesis. Although a number of transgenic RNAi constructs directed against SCAR and functionally-associated elements failed to elicit an effect, mef2-gal4-based expression of a SCAR-RNAi construct from the potent VALIUM series [89, 90] strongly disrupted flight muscle development, generating a characteristic myoblast fusion-arrest phenotype of unfused myoblasts congregated around thin, under-developed DLM templates (Fig 6D-E ). Such myogenic defects (Fig. 6F-F ) were also observed following similar utilization of a VALIUM-series RNAi construct targeting kette, which encodes the conserved NAP1 component of the SCAR regulatory complex [73]. Zygotic loss-offunction mutations in this element result in a strong arrest of embryonic myoblast fusion, and have been used as a major tool for study of SCAR function in this context [20, 68]. Specificity of the kette RNAi construct was verified by restoration of normal myogenesis following co-expression of UAS-kette (see Fig. 6 legend). As expected, strong fusionarrest phenotypes were observed following RNAi-based targeting of SOP2 and Arp2 [91], two subunits of the Arp2/3 complex (Fig. 6G-H ), the downstream target of both the Wsp and SCAR NPFs. Quantification of DLM nuclear content (Fig. 6J) underscores the strong inhibition of myoblast fusion by RNAi targeting the Arp2/3 complex, or either of its major NPF systems. 47
 fail to grow and split following expression of UAS-RNAi targeted against Wsp (B) and D-WIP (C), via mef2-gal4.")
48 Fig. 6. Muscle-specific RNAi expression reveals a requirement for the SCAR NPF in adult myoblast fusion. (A-C) Dissected DLMs at 24hrs APF, visualized with Mab 22C10. DLM templates (marked with stars) fail to grow and split following expression of UAS-RNAi targeted against Wsp (B) and D-WIP (C), via mef2-gal4. (D-H ) DLMs at 24hrs APF, following mef2-gal4-based expression of transgenic UAS- RNAi constructs. Fibers (green) are visualized with Mab22C10, and nuclei (red) with anti-ewg. Ewg is expressed in all nuclei within the fibers, as well as in nuclei of surrounding myoblasts. The wild-type (WT) panels (D-D ) represent the driver on its own. RNAi constructs used were targeted against SCAR (E-E ), kette (F-F ), SOP2 (G-G ) and Arp2 (H-H ). Scale bar = 50 µm (A). (I) The extent of DLM fusion was quantified by counting Ewg-positive nuclei within single DLM fibers at 24hrs APF. Wild-type fibers contain nuclei (n=4). Quantification of Ewg-positive nuclei within DLMs of Wsp 1 /Df(3R)3450 mutant pupae serves as the measure of fiber nuclear content in the complete absence of fusion (see also Fig. 3D and accompanying text). Fusion is strongly inhibited following expression in muscles of D-WIP-RNAi ( nuclei, n=10), Wsp-RNAi ( nuclei, n=8), SCAR-RNAi ( , n=10), kette-rnai ( , n=10), Arp2-RNAi ( , n=9) and SOP2-RNAi ( , n=6). Co-expression of UASkette (Hummel et al., 2000) together with the kette RNAi construct significantly rescues the fusion defect ( nuclei, n=9), verifying specificity of the RNAi construct. The minimal degree of fusion observed for all RNAi transgenes, including the Wsp-targeting construct, attests to the difficulty of obtaining complete disruption of gene activity by this approach. 48
49 Arp2/3, but neither of its major NPFs, mediates formation of fusion- associated actin foci The demonstrated involvement of actin polymerizing factors during adult myoblast fusion implies the formation of microfilament structures that contribute to the fusion process. To try and identify such structures, we monitored the sub-cellular distribution of phalloidin-stained microfilaments during DLM formation. The analysis revealed that single, prominent, spherical F-actin structures form at the interface between DLM fibers and attached myoblasts, on the verge of fusion (Fig. 7A). We refer to these structures as actin foci, a term used to describe F-actin structures of similar appearance and size (2-3 um 2 in area), that are associated with myoblast fusion during Drosophila embryogenesis [34, 68, 75]. Such foci are also observed during construction of the adult de novo forming DVMs (Fig. S3), suggesting that they are a general feature of myoblast fusion in Drosophila. To assess the manner in which the foci are distributed between DLM fibers and fusing myoblasts, we visualized them following cell-type specific expression of the GFP-tagged F-actin binding domain of Moesin [92, 93]. Full overlap (Fig. 7B-B ) was commonly observed between 1151-GAL4-expressed Moe-GFP and phalloidin (19/20 foci examined), while overlap of rp298-gal4-expressed Moe-GFP and phalloidin was rarely seen (2/18 foci), and even then was restricted to a minimal portion of the structure (Fig. 7C-C ). The strong bias obtained using this approach suggests therefore, that the actin foci reside primarily, if not exclusively, within myoblasts. We next sought to determine whether the Arp2/3 complex and its associated NPFs contribute to formation of the fusion-associated actin foci. Towards this end, we first monitored the localization pattern of two subunits of the Arp2/3 complex, Arp3 and SOP2/Arpc1, by expression of GFP-tagged versions [91] in fusing adult myoblasts. Both constructs were found to strongly co-localize with the actin foci, consistent with a role for Arp2/3 in their establishment (Fig 7D-E ). Indeed, following expression of RNAi constructs targeted against the Arp2/3 subunits Arp2 and SOP2 via mef2-gal4, the bright, spherical foci were replaced by small and diffuse accumulations of actin (Fig. 7G- I), implying that Arp2/3 function is essential for properly constructing the fusionassociated foci. 49
50 We next used RNAi-based disruption of gene activity to determine the identity of the NPF system that stimulates Arp2/3 activity in this context (Fig. 7J-P). Although D- WIP co-localizes with the actin foci (Figure 7F), expression of RNAi constructs targeting Wsp (Fig. 7J) and D-WIP (Fig. 7K) did not interfere with the capacity to form foci of normal size. Turning to the SCAR-based NPF system, we observed that expression of RNAi constructs targeting either SCAR or the SCAR-complex element kette, led to formation of enlarged, irregularly-shaped actin foci (Fig. 7L,M). Enlarged foci are similarly generated following simultaneous RNAi-based targeting of both the Wsp and SCAR-based NPF systems (Fig. 7N,O). Thus, neither separate nor combined Wsp and SCAR-complex NPF function appear to directly mediate the contribution of the Arp2/3 complex to actin focus construction, implying the involvement of a distinct, unknown system for stimulation of Arp2/3 actin nucleation activity in this context. As discussed below, the nature of the involvement of the WASp and SCAR/WAVE pathways in actin focus formation in adult myoblasts, mirrors observations made during myoblast fusion in Drosophila embryos. The ability to directly monitor Arp2/3 complex function is, however, unique to the adult system, and leads us to suggest that the fusion-associated foci are Arp2/3-dependent branched microfilament structures, and that a novel NPF mediates the involvement of Arp2/3 in this process. 50
 Sphere-shaped foci containing F-actin (arrows) are present at the interface between DLM templates (visualized with Mab22C10, green) and attached myoblasts, in an 18hrs APF pupa.")
51 Fig. 7. Arp2/3-dependent F-actin foci form during adult myoblast fusion. (A) Sphere-shaped foci containing F-actin (arrows) are present at the interface between DLM templates (visualized with Mab22C10, green) and attached myoblasts, in an 18hrs APF pupa. Foci and myoblast outlines are visualized with phalloidin (red). (B-B ) An 18 hrs APF pupa expressing the F-actin binding protein Moe- GFP, under control of the myoblast-specific driver 1151-GAL4. The anti-gfp (B, green) and phalloidin (B, red) staining patterns completely overlap (B, merge). In contrast, when expressing Moe-GFP via the fiber-specific driver rp298 (duf)-gal4, the anti-gfp (C, green) and phalloidin (C, red) staining patterns 51
52 are mostly separate (C, merge), in the rare instances where overlap is detected altogether. The actin focus therefore resides primarily, if not exclusively, within myoblasts. (D-E ) 18 hrs APF pupae expressing GFPtagged versions of the Arp2/3 subunits Arp3 (D-D ) and SOP2 (E-E ). Both subunits, visualized using anti- GFP (green), strongly co-localize with F-actin (phalloidin, red) at focus structures (arrows in merged panels). (F-F ) Endogenous D-WIP in an 18 hrs APF pupa, visualized with anti-d-wip antibodies (green), is also found to co-localize with the F-actin (phalloidin, red) focal structures (arrows in merged panel). (G- O) Morphology of 18hr APF actin foci (visualized with phalloidin, red) following RNAi-based targeting of Arp2/3 and related NPFs, via the mef2-gal4 driver. DLM template muscles are visualized with Mab22C10 (green) in all panels. (G) Wild-type sized focus in the absence of RNAi construct expression. Weak, diffuse foci are observed following targeting of the Arp2/3 subunits Arp2 (H) and SOP2 (I). Targeting of Wsp (J) and D-WIP (K) does not affect focus size and morphology, while enlarged, somewhat irregular foci are found in pupae expressing RNAi targeting SCAR (L), the SCAR complex element kette (M) and both Wsp and SCAR (N) or Wsp and kette (O). (P) Bar graph displaying measured areas of actin foci in pupae expressing different RNAi constructs. Values represent average area of ~25 foci for each genotype. Asterisks indicate foci size values that differ significantly (P<0.005) from wild-type values. Scale bars= 5 µm (A), 2 µm (B,D,G). 52
53 3.3. Discussion Involvement of the Arp2/3 nucleation system in formation of fusion-associated actin foci Spherical, F-actin rich structures, closely associated with myoblast fusion pore formation during Drosophila embryogenesis, have been recently described and extensively characterized [68, 76, 94]. Our observation of highly similar structures during the period of adult muscle growth via fusion, now suggests that these foci are a universal feature of the myoblast fusion process. While a variety of alterations in embryonic actin focus morphology have been described for various mutants, the molecular mechanism governing their formation is still unknown. A key, lingering question is the identity of the actin nucleation machinery responsible for establishment of the foci. The Arp2/3 nucleation system is an obvious candidate for such a role, given the well-documented involvement of its different elements in embryonic myoblast fusion. However, direct assessment of Arp2/3 complex function during embryogenesis is difficult, since the maternal contribution of complex subunits, which is essential for oogenesis [66, 91], is sufficient to overcome zygotic gene disruption. The capacity of RNAi to efficiently disrupt gene function during adult myogenesis thus provides a unique opportunity to study the contribution of the Arp2/3 complex to actin focus formation. Our data suggests that Arp2/3 indeed plays a critical role in this process, as focus formation is significantly impaired upon expression of RNAi targeting two of the complex subunits. Such a requirement for Arp2/3 is somewhat surprising, since, as shown above, actin foci persist or grow following separate or simultaneous disruption of the two major Arp2/3 NPF systems, centered on Wsp and SCAR. Furthermore, these behaviors closely match those reported following disruption of Wsp and SCAR system activity, by a variety of means, during embryogenesis [68, 76, 86]. While the WASp and SCAR/WAVE systems continue to be regarded as primary NPFs for Arp2/3, a growing list of alternative nucleators has recently emerged [95], suggesting that a novel element may well activate Arp2/3 in this particular context. Identification of this novel NPF should provide genetic tools for determining the functional significance of the actin foci, 53
54 which has so far remained elusive, due to the lack of solid knowledge regarding the mechanism by which they are constructed. In summary, the observations reported in our study generalize the involvement of the Arp2/3 complex and its associated cellular machinery during construction of muscle fibers via myoblast fusion. In particular, a prominent, and potentially universal role is assigned to the WASp NPF as a mediator of these events. 54
55 4. Fusion competence is repressed in migrating Drosophila myoblasts by Notch signaling 4.1. Introduction Multi-nucleated muscle fibers are characteristic of the musculature of most multicellular organisms, accommodating the need to generate large and continuous fibers. Generation of myofibers from individual myoblasts has been studied in great detail in the Drosophila embryo, where analysis of mutants has allowed the identification of key players. This is a complex process, involving the specification of cells that will serve as founders for each of the muscles, the definition of myoblasts, and the attraction of myoblasts towards founder cells. Once the two cell types are in close contact, cell fusion is executed via an elaborate machinery involving cell adhesion, formation of fusion pores between the attached cells and dissolution of cell membranes [96]. Which aspects of this process are unique to the Drosophila embryo, and what mechanisms will turn out to be common to all muscle fusion processes? One may expect the specification of distinct muscle cells to be more divergent for different organisms, while the actual machinery of fusion is likely to be more universal in nature [97-99]. The somatic musculature of Drosophila embryos is formed from two subpopulations of myogenic mesoderm myoblasts, founder cells and the more numerous fusion-competent myoblasts (FCMs). Each of the thirty-odd muscles of an embryonic hemi-segment arises from a single, specific founder cell, that expresses a distinct combination of transcription factors. Sequential rounds of fusion of each founder cell with neighboring FCMs generates myofibers of characteristic shape and size, in which all nuclei assume the specific muscle fate of the original founder cell. The overall number of myoblasts that are initially defined within the myogenic mesoderm is sufficient to provide the necessary muscle mass, even without additional cell divisions prior to fusion. In addition, some myoblasts are set aside as individual adult progenitor cells, and will provide the source of cells for the adult muscles [100]. The cell adhesion molecule Dumbfounded (Duf) is expressed exclusively in the founder cells. Duf has been suggested to provide an attractive cue for the FCMs, thereby mediating founder cell-fcm recognition and attachment [10]. Duf likely performs this 55
56 function together with its paralouge Roughest (Rst) [11], whose expression pattern is less restricted. Conversely, the Ig-domain proteins Sticks and stones (Sns) and Hibris (Hbs) are expressed by the FCMs, and through heterotypic interactions with Duf/Roughest form the initial contact between the two cell types [12-15]. Following Duf/Sns association, the fusion process ensues. A large number of elements mediating myoblast fusion in Drosophila embryos has been identified (1). Many of these are known to function in the context of the actin-based cytoskeleton, and indeed a large, transient actin focus structure is present at sites of myoblast fusion [16, 34, 68, 86, 101]. Ultra-structural analysis has given rise to a number of models for progress of fusion at the sub-cellular level, in which roles for cell adhesion, vesicular entities and cytoskeletal structures have been proposed to provide a mechanistic basis for fusion pore formation and plasma membrane dissolution [34-36, 101, 102]. The process of multinucleated muscle formation is recapitulated during the pupal stages of Drosophila development, when nearly the entire larval musculature undergoes histolysis, and new adult muscles are generated [103]. Similar to the embryo, most multi-nucleated adult muscle fibers are seeded by a founder cell, which fuses with neighboring myoblasts. An alternative myogenic program makes use of a few subsets of larval muscles that escape histolysis, and then provide templates for formation of adult muscles. Prominent among these is the process that generates the dorsal longitudinal indirect flight muscles (DLMs), a set of twelve large muscles housed within the adult fly thorax. In this case, a population of adult myoblasts positioned on the wing disc, migrate during early pupal stages towards a group of persistent thoracic larval muscles and fuse with them, leading to a significant enlargement of muscle mass [1]. Before and during the course of migration, these myoblasts divide, and can thus be regarded as transit amplifying cells, prior to their fusion with the myotube templates. Another unique feature of these cells is their migration as a swarm of closely associated cells, throughout their transit from the wing disc to the muscle templates [2]. It should also be noted that wing morphogenesis and other tissue movements in the developing pupa, generate a highly dynamic landscape in which these cells migrate. The underlying external cues which guide the migration of myoblasts, and the putative signals they may transmit to each other during the swarming process, to maintain their transit-amplifying state, are not known. 56
57 Analysis of myogenesis during pupal stages has been limited by the difficulties in carrying out mutant clone analysis in a multinucleated tissue. We have previously shown that an RNAi-based approach can overcome these difficulties, in demonstrating the crucial involvement of Wsp, the sole fly homolog of WASP-family actin nucleationpromoting factors, as well as other actin regulators, during adult myoblast fusion [82]. Here we extend this approach to study the myogenic events preceding onset of fusion in DLM muscles, including attraction and migration of myoblasts towards the DLM muscle templates, and the program of transcriptional regulation that endows myoblasts with a competence to fuse. In particular, we show that Duf is required for attraction of migrating wing disc-derived myoblasts to DLM templates, as well as for the initial contact between myoblasts and myotubes. During migration as a swarm, we find that the myoblasts are maintained in the semi-differentiated, proliferating state, by continuous activation of Notch pathway signaling, where each myoblast provides the ligand Delta (Dl), to its neighbors. Notch activity within the semi-differentiated myoblasts inhibits the ability of myoblasts to fuse by repressing expression of key fusion-related factors such as Sns and the actin regulator D-WIP. Notch signaling between myoblasts decays once they reach the vicinity of the DLM myotubes, leading to terminal differentiation of the myoblasts, and triggering the onset of fusion by induction of sns expression. 57
58 4.2. Results Duf provides an attractive cue to migrating imaginal myoblasts During embryonic myogenesis Duf is expressed exclusively in founder cells and myotubes, and the protein is detected on the plasma membrane [10, 104]. Duf expression reporters have suggested a role for the protein during pupal stages as well [105]. In thoracic pupal preparations during metamorphosis, we detected the Duf protein mainly on the membrane of the developing DLM myotubes from hours after puparium formation (APF), as well as in intracellular puncta [Figure 1A]. Duf localization on the myotube membrane is not uniform, but rather is enriched at contact points with fusing myoblasts [Figure 1A''], as can be ascertained by co-localization with D-WIP, which marks myotube-myoblast contact sites [17]. Disruption of both Duf and Rst function was achieved by simultaneous expression of RNAi constructs in myogenic tissue (via mef2-gal4), in combination with the hypomorphic duf rp298 allele. In this genetic background, where Duf protein is no longer detectable [Figure 1B-B ], myoblast behavior shows considerable abnormalities. Examination of thoracic muscles reveals that most myoblasts do not reach the myotubes when Duf and Rst function is compromised in this manner [Figure 1D,G compare to Figure 1C,F]. The Duf/Rst mutant phenotype is dramatically different from that observed when myoblast fusion to the DLM templates is disrupted, as seen in Wsp mutants or following Wsp knockdown via RNAi [82]. When fusion is halted in this manner, myoblasts readily reach the DLM myotubes, and create a cloud of unfused, swarming myoblasts around them [Figure 1E,H]. In Duf/Rst knockdown pupae, although myoblasts are properly positioned on the wing imaginal discs at earlier stages of development (Supplementary Figure 1), they scatter throughout the pupa instead of migrating towards the myotubes. This result indicates that expression of Duf and Rst in the myotubes is required for attracting the migrating myoblasts. Next, we sought to investigate if Duf is also sufficient for myoblast attraction. Towards this end, Duf was ectopically expressed in the dorsal epidermis of the thorax (notum) using the ND637-GAL4 driver [106]. The spatial pattern of wing disc-derived myoblasts was monitored by visualizing the m6-gfp Notch reporter, which marks the myoblasts from larval stages, during migration, and up until the onset of fusion with the 58
59 DLM templates [107]. Following ectopic Duf expression in a wild-type pupa, a dramatic alteration in myoblast distribution was observed. While some myoblasts migrate normally to surround the myotubes, a significant fraction is drawn to the ectopic Duf expression zone in the epidermis, and the cells spread directly underneath this domain [Figure 2B, compare to 2A]. 59
![Figure 1. Duf is expressed on myotubes and attracts myoblasts. [A,B] Dissected pupal muscle preparations, 18-22 hours APF.](/docs-images/104/165733799/images/60-0.jpg "[A,A'] Staining for the Duf protein (white, red) in control flies (sns- LacZ/CyO), demonstrating its specific expression only in myotubes but not in myoblasts, which express either the nuclear marker")
60 Figure 1. Duf is expressed on myotubes and attracts myoblasts. [A,B] Dissected pupal muscle preparations, hours APF. [A,A'] Staining for the Duf protein (white, red) in control flies (sns- LacZ/CyO), demonstrating its specific expression only in myotubes but not in myoblasts, which express either the nuclear marker Twi (green) or D-WIP (blue), which is up-regulated in myoblasts prior to fusion 60
![(17). Duf is enriched in myotube membrane domains adjacent to D-WIP expressing myoblasts (white arrows in [A ] and enlargement in [A'']).](/docs-images/104/165733799/images/61-0.jpg "[B, B', enlargement in B''] Efficient elimination of the Duf protein was achieved by combining a hypomorphic duf allele with expression of duf RNAi constructs (duf rp298/y ; duf RNAi,rst RNAi /+;")
61 (17). Duf is enriched in myotube membrane domains adjacent to D-WIP expressing myoblasts (white arrows in [A ] and enlargement in [A'']). [B, B', enlargement in B''] Efficient elimination of the Duf protein was achieved by combining a hypomorphic duf allele with expression of duf RNAi constructs (duf rp298/y ; duf RNAi,rst RNAi /+; mef2-gal4, UAS-CD8GFP,UAS-Dcr2/+). [C-E] Thoraces of live pupae hours APF. Images are projections of µm thick Z-stacks. DLM templates are indicated by green arrows [F-H] Dissected pupal muscle preparations, hours APF. Myotubes are indicted by green outlines in the insets. Myoblasts and myotubes in all panels (C-H) are visualized by GFP. [C,F] Control flies (mef2- GAL4,UAS-CD8GFP,UAS-Dcr2/+) display growing sets of DLM myofibers (indicated by green arrows), surrounded by numerous myoblasts, visualized by GFP, migrate and fuse to the muscle temples. [D,G] In flies in which Duf activity is disrupted (as in panels B-B ) only a small number of myoblasts reach the templates, which remain thin and small. [E,H] In Wsp knockdown pupae (mef2-gal4,uas-cd8gfp,uas- Dcr2/ UAS-Wsp RNAi ), myoblasts aggregate around the templates but fail to fuse with them. Scale bars in [A- B, F-H] are 20µm, in [A'', B''] 5µm and in [C-E] 100µm. Figure 2. Duf is sufficient to attract myoblasts. [A,B] Live imaging of myoblasts with the m6-gfp Notch reporter (green). Notal tissue (red) is visualized by expression of myristoylated RFP via the epidermal ND 637 -GAL4 driver. Images are projections of 70-85µm thick Z-stacks. [A] In control pupae (UAS-myrRFP/+ ; ND 637 -GAL4,m6-GFP/+) at 18 hours APF, the myoblasts migrate ventrally and surround the DLM muscle templates. [B] When Duf was ectopically expressed in the epidermis (UASmyrRFP/UAS-Duf ; ND 637 -GAL4,m6-GFP/+), the myoblasts changed their spreading pattern, and a significant portion accumulated directly underneath the epidermal Duf-expressing cells. [A',B'] Higher resolution images of the X-Z plane (broken lines in [A,B]), showing the close proximity of myoblasts to the epidermal cells when the latter express Duf. Scale bars in [A-B] are 100µm and in [A'-B'] 20µm. Duf is required for fusion To ascertain whether Duf is required during the fusion process itself, we made use of UAS-FlagTM-CT [108], a putative dominant-negative Duf construct, in which nearly the entire extracellular domain of the protein is missing, while the signal peptide, transmembrane and cytoplasmic domains are retained. Restricted expression of this construct in myotubes (using rp298-gal4, a GAL4 insertion in the duf locus), did not interfere with the function of wild-type Duf as an attractant, as myoblast migration 61
62 towards the DLM templates was not impeded. However, myoblast fusion with the myotubes was strongly attenuated, evidenced by the accumulation of unfused myoblasts, abnormal splitting of the templates and aberrant muscle morphology (Supplementary Figure 2). Taken together, these results indicate that Duf is a key player in adult myogenesis. While specifically expressed in myotubes, Duf has a critical role in directing myoblast migration towards the myotubes, and a subsequent, essential role in mediating fusion, presumably by promoting adhesion between myoblasts and myotubes. Sns/Hbs function mediates establishment of myoblast-myotube fusion sites Heterotypic interactions between the Duf/Rst and Sns/Hbs pairs of cell-surface proteins is generally accepted as the molecular basis for association between founder cells/myotubes and myoblasts in Drosophila embryos. sns mutant embryos display phenotypes very similar to duf, rst double mutants: compromised migration of the myoblasts, and absence of adhesion between myoblasts and founder cells/myotubes [12, 109]. Enhancement of these phenotypes, leading to a complete fusion block, is observed in sns, hbs double mutant embryos [15, 108]. Separate knock down of sns or hbs in the pupa using RNAi constructs did not affect flight muscle formation (not shown), but simultaneous knock-down of both elements resulted in a strong fusion arrest. However, in contrast to the dispersal of myoblasts observed following duf, rst knockdown, myoblasts in the sns hbs double knockdown had clearly reached the DLM templates and accumulated around them [Figure 3B]. Further examination suggests that this phenotype results from a block of the fusion process at the initial phase of myoblast/myotube contact. Distinct actin foci have been shown to form at sites of myoblast fusion during adult muscle formation, and persist in mutants such as Wsp, where fusion arrests after myotube/myoblast contact is established [82]. Indeed, we observed numerous actin foci at the interface between DLM template myotubes and myoblasts following RNAi-based Wsp knockdown [Figure 3C], but such foci were completely absent from sns, hbs knockdown pupae [Figure 3D]. Thus, following sns, hbs knockdown migrating myoblasts reach the DLM myotubes, but fusion sites are not initiated. The duf,rst and sns,hbs double knockdown phenotypes lead us to propose a sequence of events enabling fusion between wing disc-derived myoblasts and the DLM 62
63 template myotubes. The process begins with the attraction of myoblasts to the templates through Duf and Rst activity, and continues by myoblast-myotube attachment and establishment of fusion sites, mediated by Sns/Hbs-Duf/Rst interactions. These events set the stage for execution of myoblast-myotube fusion, which will involve creation of F-actin foci at the fusion sites, fusion pore formation and membrane breakdown. 63
![Figure 3. Sns and Hbs are required for myoblast fusion and formation of actin foci. [A-D] Dissected pupal muscles 18-22 hours APF.](/docs-images/104/165733799/images/64-0.jpg "[A-B] GFP (green) was driven by mef2-gal4 to mark both myoblasts and myotubes, and generate a muscle restricted gene knockdown.")
64 Figure 3. Sns and Hbs are required for myoblast fusion and formation of actin foci. [A-D] Dissected pupal muscles hours APF. [A-B] GFP (green) was driven by mef2-gal4 to mark both myoblasts and myotubes, and generate a muscle restricted gene knockdown. Twi (red) stains only myoblasts, while Duf 64
65 (blue) is specific to myotubes. [A] In control flies (mef2-gal4,uas-cd8gfp, UAS-Dicer2/+), where fusion occurs normally, the three DLM larval templates in each hemi-thorax grow in size and split, generating a set of six thick muscle fibers. [B] When both sns and hbs are knocked down at the 3 rd instar larvae stage (GAL80 ts /+; mef2-gal4,uas-cd8gfp,uas-dicer2/sns RNAi,hbs RNA i ), the myoblasts completely fail to fuse with the DLM templates. As a result, Twi-positive myoblasts remain clustered around the thin templates, that also fail to split. [C-D] Phalloidin (white, blue) was used to follow the formation of F-actin foci at the myotube/myoblast interface. GFP (green) marks all the myogenic cells.[c] When fusion is blocked by Wsp-RNAi (mef2-gal4,uas-cd8gfp, UAS-Dicer2/Wsp RNAi ), numerous actin foci are observed, marking the sites of fusion arrest. In contrast, when fusion is blocked by sns,hbs RNAi (GAL80 TS /+; mef2-gal4,uas-cd8gfp,uas-dicer2/sns RNAi,hbs RNA i) no actin foci are formed. Scale bars are 20µm in [A-D] and 5µm in [C'', D'']. Sns is expressed in terminally differentiated adult myoblasts The pupal sns,hbs double knockdown phenotype differs from the corresponding embryonic mutant phenotypes, in that adult myoblasts appear capable of recognition of an attractive signal and migration towards myotubes. To address the basis for this more restricted role of the Sns/Hbs adhesion proteins, we monitored the expression pattern of sns during DLM growth, using the expression reporter sns-lacz [110]. Reporter expression, determined using anti-β-gal antibodies, revealed that sns expression is restricted to a small subset of myoblasts positioned near the myotubes, but is absent from the bulk of the myoblast population (visualized using anti-twist (Twi)) [Figure 4A]. This is in sharp contrast to the situation in the embryo, where sns expression is detected in all fusion-competent myoblasts [12]. In addition to the limited myoblast expression pattern, sns-lacz was detected in all DLM myotubes. This appears, however, to be an indirect consequence of incorporation of sns-lacz expressing myoblasts into the templates, rather than a reflection of de novo myotube expression. Thus, β-gal staining is completely absent from myotubes of Wsp knockdown pupae, in which fusion is blocked [Figure 3C]. The position of the sns-lacz expressing cells in close vicinity to the myotubes, suggested that they represent a late step in the gene-expression program of myoblasts. Indications of a distinct late-stage expression program are provided by the temporal expression pattern of Twi and D-WIP. Twi is expressed in the majority of myoblasts, yet is down regulated in myoblasts just prior to fusion [100, 111], while D-WIP is upregulated just before the formation of the actin foci and fusion [82]. Indeed, the subpopulation of sns-lacz expressing myoblasts is mostly Twi negative (~90%) and D-WIP positive (~70%) [Figure 4A, C]. 65

66 Taken together, these observations imply a dynamic transcriptional program in wing disc-derived myoblasts, in which mature myoblasts lose some of their early markers, while initiating expression of elements necessary for fusion with the DLM templates. sns transcription is limited to this sub-population of myoblasts. Such restriction and late onset of sns expression within the myoblast population can explain why sns function is dispensable for myoblast migration, and is consistent with a functional role during myoblast-myotube attachment. Figure 4. Sns is expressed in a small subset of myoblasts. [A,B,D] Dissected pupal muscles hours APF. [A] The Sns reporter sns-lacz reveals that Sns (green) is expressed in myoblasts immediately adjacent to myotubes, and is co-expressed with D-WIP (blue) but not with Twi (red). Duf (magenta) marks the myotubes. [B] When fusion is blocked by using Wsp RNAi (GAL80 TS /sns-lacz; mef2-gal4,uas- CD8GFP,UAS-Dicer2/Wsp RNAi ), expression of Sns (green) is observed in Twi (red) negative myoblasts, but not in myotubes marked by Duf (magenta). This implies that sns-lacz expression is restricted to myoblasts, and that its presence in myotubes [A-A ] is a consequence of fusion. [C] Quantification of the percentage of Sns-expressing myoblasts that co-express Twi (red, 9.3%±5.8, three different experiments, 40 cells per experiment), Notch reporter m6-gfp (green, 29.6%±4.7, three different experiments, 30 cells per experiment) and D-WIP (blue, 71.6%±4.4, three different experiments, 40 cells per experiment). [D,D'] Flies carrying both sns-lacz (red) and the Notch reporter m6-gfp (green) display mutually exclusive expression patterns, and a positive correlation between the Notch reporter and Twi (blue). Scale bars in [A- D] are 20µm. Notch signaling maintains the transit-amplifying state Notch signaling has been implicated in the context of adult myogenesis in Drosophila [ ]. An interesting feature is that Notch signaling is active in most wing disc-derived myoblasts, but is shut down in a small number of these myoblasts that are in close proximity to the myotubes [112]. Using the Notch reporter line m6-gfp 66
67 [107], we found that the myoblasts displaying reduced Notch signaling also show a decline in Twi levels [[112] and Figure 4D]. To test if sns expression is linked to a reduction in Notch signaling, we examined sns-lacz and m6-gfp expression in myoblasts of pupae harboring both reporters. Our analysis revealed that in most cases, cells that up regulate Sns, down regulated m6-gfp (~70%) [Figure 4D, quantified in 4C]. Perdurance of GFP induced by previous Notch activation is likely to make this an underestimate of the proportion of Sns-expressing cells in which Notch signaling is no longer active. To investigate whether Sns repression is under control of Notch, we chose to manipulate Notch signaling and monitor both myoblast fusion and the expression of Sns. We first expressed the constitutively active intracellular domain of Notch (N ICD ) [114] in both myotubes and myoblasts, under the potent mef2-gal4 driver. This resulted in a striking phenotype: The expression of sns was completely absent from myoblasts, and fusion between myoblasts and DLM templates was blocked [Figure 5A]. Next, we conducted the complementary experiment, generating a Notch loss-offunction specifically in the myogenic lineage. This was most efficiently obtained by expressing Notch RNAi via the mef2-gal4 driver. The results were also dramatic: Not only were Sns expressed and Twi repressed in all myoblasts, but the size of the unfused myoblast population was strikingly small [Figure 5B, additional time points in Supplementary Figure 3], suggestive of premature myoblast differentiation and an accelerated rate of myoblast-myotube fusion. To expand the size of the myoblast pool under conditions of Notch knockdown, we blocked fusion by simultaneous knockdown of sns and hbs. The triple knockdown resulted in a full arrest of fusion, and essentially all myoblasts were positive for Sns and negative for Twi expression. Taken together, the Notch gain- and loss-of-function experiments imply that Notch signaling suppresses the expression of Sns. Only in mature myoblasts that are in close proximity to the myotube does a decline of Notch signaling allows induction of Sns expression, just before the onset of fusion. 67
![Figure 5. Notch signaling represses Sns expression and myoblast fusion. [A-C] Dissected pupal muscles 18 hours APF.](/docs-images/104/165733799/images/68-0.jpg "All crosses were carried out in the Gal80 TS background, and the temperature shift was done at the wandering larvae stage, to induce the UAS constructs only during metamorphosis.")
68 Figure 5. Notch signaling represses Sns expression and myoblast fusion. [A-C] Dissected pupal muscles 18 hours APF. All crosses were carried out in the Gal80 TS background, and the temperature shift was done at the wandering larvae stage, to induce the UAS constructs only during metamorphosis. [A] In flies in which the Notch pathway is costituitively active (GAL80 ts /sns-lacz,uas-n ICD ; mef2-gal4,uas- CD8GFP,UAS-Dicer2/+), accumulation of Twi-positive myoblasts (blue), and absence of myoblast fusion leading to thin myotubes (marked by Duf (red)) was observed. Expression of sns-lacz (green) was completely blocked in myoblasts, but maintained in other tissues. [B] Notch knockdown (UAS-N RNAi /+; GAL80 TS /sns-lacz; mef2-gal4,uas-cd8gfp,uas-dicer2/+) caused premature differentiation marked by Sns induction and Twi repression, while fusion proceeded readily. [C] When both Notch and sns/hbs are knocked down (UAS-N RNAi /+; GAL80 TS /sns-lacz; mef2-gal4,uas-cd8gfp,uas-dicer2/ sns RNAi,hbs RNA i ), Sns-positive/Twi-negative myoblasts accumulated around the myotubes, but did not fuse with them. Scale bars in [A-C] are 20µm. 68
69 Delta-Notch signaling between myoblasts prevents premature differentiation Notch signaling has been intensively studied in epithelial tissues, where the cells are stationary, and ligand-receptor interactions occur between permanent neighboring cells (reviewed in [115]). Migrating mesenchymal cells pose a challenge for canonical Notch signaling. Such cells can associate with a variety of neighbors along the way, and constantly alter their morphology. Thus, the dynamics of ligand sensing should adapt to these conditions. Since wing disc-derived pupal myoblasts migrate as a swarm, other cells in the swarm represent an obvious option for ligand presentation by neighboring cells. We stained the swarming myoblasts and developing myotubes for the two Notch ligands Dl and Serrate (Ser). Both ligands are presented on the myogenic cell membranes. Ser has a distinct discontinuous membrane pattern, mostly restricted to the myoblasts [Figure 6A], while Dl shows a general membranal staining in both myoblasts and myotubes [Figure 6B]. We sought to use the RNAi-based knockdown approach in order to assess the function of the two Notch ligands. Separate expression of RNAi constructs using the mef2-gal4 driver, resulted in pronounced reductions in ligand epitope detection for both Ser [Figure 6C] and Dl [Figure 6D]. These observations demonstrate the capacity to interfere with Notch ligand function, and furthermore, attest to the specificity of the antibodies used for ligand localization. Reducing the levels of Ser by RNAi-knockdown did not result in a prominent phenotype at 20 hours APF, i.e. the number of Twi-expressing myoblasts was comparable to age matched wild-type pupae [Figure 6F]. However, reducing the levels of Dl gave rise to a dramatic phenotype: Unfused myoblast number was abnormally low, and the few that did not fuse were Sns positive and Twi negative [Figure 6G]. While the Dl knockdown phenotype is similar to that of Notch knockdown [Figure 6E], the Notch knockdown phenotype is more severe. At this phase there are no unfused myoblasts in Notch knockdown pupae, and the DLM muscles are unsplit and smaller. Taken together, these observations suggest that in the myoblast swarm, Notch activation levels are maintained predominantly via Dl-Notch interactions between the myoblasts. 69
![Figure 6. Delta is the prominent ligand that signals to Notch in imaginal myoblasts. [A-D] Dissected pupal muscles 20 hours APF.](/docs-images/104/165733799/images/70-0.jpg "[A,B] The Notch ligands Ser [A] and Dl [B] are displayed on both myoblasts and myotubes in wild-type pupae. Ser appears to be enriched on myoblast membranes.")
70 Figure 6. Delta is the prominent ligand that signals to Notch in imaginal myoblasts. [A-D] Dissected pupal muscles 20 hours APF. [A,B] The Notch ligands Ser [A] and Dl [B] are displayed on both myoblasts and myotubes in wild-type pupae. Ser appears to be enriched on myoblast membranes. [C] Ser RNAi (GAL80 TS /sns-lacz,uas-ser RNAi ; mef2-gal4,uas-cd8gfp,uas-dicer2/+) can efficiently knock down Ser levels in developing DLMs. [D] Dl RNAi (GAL80 TS /sns-lacz,uas-dl RNAi ; mef2-gal4,uas- CD8GFP,UAS-Dicer2/+) can efficiently knock down Dl levels in developing DLMs. [E] In Notch RNAi pupae 24 hours APF (UAS-N RNAi /+; GAL80 TS /sns-lacz; mef2-gal4,uas-cd8gfp,uas-dicer2/+) no Twi-positive myoblasts (blue) are present, and the few remaining unfused myoblasts are Sns positive (red). [F] Pupae 24 hours APF, expressing Ser RNAi (GAL80 TS /sns-lacz,uas-ser RNAi ; mef2-gal4,uas- CD8GFP,UAS-Dicer2/+). Twi-positive cells are still present, similar to age equivalent wild-type pupae. [G] Pupae 24 hours APF, expressing Dl RNAi (GAL80 TS /sns-lacz,uas-dl RNAi ; mef2-gal4,uas- CD8GFP,UAS-Dicer2/+). In the few remaining unfused myoblasts, Twi is repressed and sns-lacz is expressed. Scale bars in [A-G] are 20µm. 70
71 A fine balance between transmitting and receiving Notch signals in myoblasts In a variety of systems where Notch signaling operates, the concomitant expression of ligand and receptor within the same cell leads to cis-inhibition, where interaction between Notch ligands and receptors leads to inactivation of both partners. This feature was suggested to play a role in reducing signaling errors under conditions where stochastic activation of Notch generates pattern [116]. Since the migrating myoblasts express both ligand and receptor, we wanted to examine the possible contribution of cis-inhibition to Notch signaling in the myoblast swarm. By generating marked flip-out clones [117], we could specifically test the effect of elevated or reduced levels of either ligand or receptor, on the capacity of a cell to receive signals from its neighbors, while leaving the ability of the adjacent cells to present the signal intact. Since myoblasts dynamically exchange neighbors in the course of migration, altering the capacity of a small number of cells to send a signal should not have a significant effect on the entire population. Flip-out clones marked by GFP expression were generated sporadically in myoblasts, by inducing a hs-flp construct at a relatively low temperature of 34 0 C. As a readout for Notch activation, we followed the capacity of the marked cells to express Twi (Notch on) or sns-lacz (Notch off). The fraction of sns-lacz-expressing myoblasts in the control flip-out cells or following co-expression of different constructs was monitored [Figure 7A]. First, we induced the expression of Notch in the flip-out cells. While this manipulation may titrate the ligand displayed by these cells, it should not compromise their capacity to receive a signal from neighboring cells. Indeed, no alteration in the fraction of sns-lacz positive cells was observed [Figure 7B]. In contrast, elevation of Notch ligand levels led to an increase in the fraction of sns-lacz-positive cells, with a more pronounced effect when Dl was over-expressed [Figure 7A,B]. These results indicate that elevation in the levels of Ser or Dl compromises the capacity of the same cells to receive a signal from neighboring cells, presumably by cis-inhibition that titrates the receptor. To monitor the consequence of a global perturbation in the balance of ligand and receptor, Ser or Dl were overexpressed in all myoblasts, using the1151-gal4 driver. Global Ser over-expression did not give rise to a marked elevation in Notch signaling, as reflected in the fraction of sns-lacz-expressing 71
72 myoblasts [Figure 7D compare to 7C]. However, Dl over-expression gave rise to a phenotype that was reminiscent of Dl or Notch knockdown (albeit weaker): a larger fraction of myoblasts (35%±10 vs. 12%±2 in wild-type) expressed sns-lacz [Figure 7E]. Thus, despite the presentation of more ligand by all cells, the predominant outcome was a reduction in Notch signaling, due to cis-inhibition of the receptor. Over expression of the N receptor in all myoblasts similarly raised the fraction of sns-lacz-expressing myoblasts [Figure 7F] (35%±8 vs. 12%±2 in wild-type), further supporting the notion that signaling can be constrained by cis-inhibition. In this case we envision that excess receptor sequesters the ligand presented by the myoblasts. Since the driver that was used is expressed only in myoblasts, this result is consistent with the notion that ligand displayed by myoblasts is the exclusive trigger of Notch activation. Over-expression in myoblast clones or in the whole myoblast population has demonstrated that higher levels of Ser or Dl can perturb the balance of Notch signaling. Are the endogenous levels of Dl and Ser fine-tuned and set at a range that is below the cis-inhibition threshold? Expression of RNAi for Ser or Dl in individual myoblast cells should allow to detect alterations in their capacity to receive the Notch signal, without altering the overall ligand-mediated signaling capacity of the myoblast population. No significant alteration in the receiving capacity of the flip-out cells was observed, when RNAi constructs for Ser or Dl were expressed [Fig. 7B]. Thus, the endogenous Ser and Dl levels are set such that they provide effective signaling between cells, but do not compromise the reception capacity. Taken together, our findings indicate that the levels of ligand and receptor co-expressed by the swarming myoblasts are finely tuned, to maintain the capacity of these cells to present and receive signals simultaneously during their migration. Proper continuous levels of Notch signaling are essential in order to sustain the swarming myoblasts in the proliferative transit amplifying state, until they encounter the DLM myotube templates. 72
.")
73 Figure 7. High levels of Ser or Delta can cis-inhibit Notch signaling. [A] A mild heat shock at 34 0 C creates a "salt and pepper" pattern of flip-out clone cells in the developing DLMs (hs-flp, UAS-GFP/+; sns-lacz/cyo; act-frt-stop-frt-gal4/+). Unfused myoblasts were monitored for the expression of GFP (green), sns-lacz (blue) and Twi (red). White arrows indicate clone cells expressing both GFP and Twi, black arrow indicates a clone cell expressing both GFP and Sns. [B] Percentage of sns-lacz positive cells out of the total number of clone cells for each experiment. For each genotype the percentage was averaged from 3-5 different experiments clone cells were counted in total for each genotype. 73
74 Asterisks denote the genotypes where a statistically-significant deviation from the wild-type ratio was observed (p-value<0.05). [C-E] Dissected pupal muscles hours APF. Images are projections of 20-40µm thick Z-stacks. [C] sns-lacz expression in a wild-type background. [D] Over-expression of Ser (1151-GAL4/+; UAS-Ser,sns-LacZ/+) did not increase the fraction of sns-lacz expressing myoblasts. [E] Over-expression of Dl (1151-GAL4/+; UAS-Dl,sns-LacZ/+) or of [F] the N receptor (1151-GAL4/+; UAS- N,sns-LacZ/+), significantly increased the fraction of sns LacZ expressing myoblasts (from 12% to 35% in both genotypes). Scale bars are 20µm. 74
75 4.3. Discussion Myogenesis in the Drosophila pupa presents a variety of challenges for faithful execution of the process, through which the adult musculature will form[118]. Here we studied the formation of the indirect DLM flight muscles, where myoblasts face the task of reaching and fusing with distant larval templates. In the course of ~20 hours of migration towards the template myotubes, these wing disc-derived myoblasts undergo continuous proliferation, in order to generate a sufficiently large pool of cells that will eventually contribute to the adult muscle mass. The myoblasts progress through two distinct phases of differentiation during the process: a semi-differentiated, amplifying phase during the course of migration; and a terminally differentiated, fusion competent phase once within close range of the myotubes. This program differs in key aspects from somatic myogenesis in the Drosophila embryo, a setting where events preceding and during myoblast fusion have been thoroughly studied. Thus, embryonic FCMs, which are to the large part post-mitotic, traverse relatively short distances towards founder cells and myotubes, and appear to behave as individual migratory cells rather than as a group [119]. Correspondingly, we find both similarities and fundamental differences in the manner by which fusion-related elements are employed and regulated in the two myogenic systems. In the current study, we show that signals from the thoracic myotubes provide the myoblasts with long-range migratory cues, while short-range signals within the myoblast swarm maintain the semi-differentiated fate of the migrating cells. In this novel and dynamic microenvironment, the myoblasts continuously exchange neighbors, yet most of the time these neighbors are identical to each other. Inhibition of the transcriptional program governing the migratory phase and onset of gene expression necessary for full differentiation and fusion, likely result from short-range, myotube-derived signals that the myoblasts encounter once they reach the vicinity of the myotubes. Our analysis of myoblast migration identifies the Ig-domain, cell-surface protein Duf, an evolutionary conserved contributor to muscle fusion in both Drosophila embryos and vertebrates [120], as a key element mediating long-range attraction of myoblasts to DLM myotube templates. As was previously shown in Drosophila embryos [10], adult myoblasts are diverted from their normal migratory path towards a source of ectopic Duf, 75
76 which can reside in a separate cell layer. However, the specific circumstances of DLM growth now characterize the Duf/Rst system as a persistent, long-range attractant for migrating myoblasts. The mechanism by which Duf/Rst provides attractive cues remains to be elucidated. Direct use of cleaved portions from the extracellular domains of these cell-surface proteins as diffusible attractants is an appealing notion. In any case, our results rule out either the cell-surface protein Sns or its paralog Hbs as myoblast receptors for the attractant, since Sns is expressed only upon completion of migration, while knockdown of hbs does not affect myoblast migration (Supplementary Figure 4). Since the normal source of Duf on myotubes is broad and myoblast attraction occurs over a long range, we do not envisage the attractive signal as a very sharp cue. In other words, it attracts the myoblast swarm to the vicinity of the myotubes, and maintains it in that general zone. Once myoblasts are at a sufficiently close range, they may encounter the myotubes stochastically. This notion is similar to the model of attraction of germ cells by Sdf1 in the zebrafish embryo, where a spatially restricted cloud of ligand maintains the germ cells within the confinement of this domain. A gradual developmental shift in the expression borders of the Sdf1 domain leads to a concomitant displacement of the germ cells towards their final target [121]. Although it has been previously shown that the myoblasts reaching the myotube change some of their molecular markers, the full significance of this process in the context of myoblast-myotube fusion has not been elucidated. Our results suggest that the myoblasts sense a continuous Notch signal, from the time the cells leave the wing disc and possibly even earlier, until the moment they reach the myotube vicinity. This signal appears to have a dual role, maintaining the cells in a proliferative mode, as well as preventing them from obtaining the competence to fuse. Notch suppression of Sns and other fusion-related genes such as D-WIP therefore prevents the untimely development of muscles. Simultaneous suppression of a battery of fusion-related genes by Notch is corroborated by our inability to rescue the Notch knockdown phenotype by ectopic expression of Sns alone (Supplementary Figure 5). Many features of pupal myogenesis resemble the repair of injured adult muscles in vertebrates. Stem cells, termed satellite cells, are located within the muscle in close proximity to the basal lamina. They undergo an asymmetric division, where one progeny 76
77 leaves its original site and undergoes proliferation, to generate a transit-amplifying population. These dividing, semi-differentiated cells do not display the full expression profile of mature myoblasts. The proliferative phase requires continuous activation of Notch, and relieves the stem cells from the need to divide asymmetrically at a mode where only a single differentiated myoblast is created at a time. The cells gradually shift to the fully-differentiated fate, first in only one progeny of each division, and eventually in all cells. The differentiated cells are capable of fusing with the injured muscle to increase its mass (reviewed in [122]). We suggest that, by analogy, the swarming Drosophila pupal myoblasts that are marked by the Notch reporter and Twi expression possess a transit-amplifying character, not yet ready to undergo fusion to the myotubes. Notch signaling in migrating pupal myoblasts is unique, in the sense that the same cell simultaneously sends and receives the signal, over a prolonged time interval. The stochastic contact of the myoblasts with each other during migration may allow them to undergo repeated cycles of Notch activation, each time triggered by exposure to a different ligand-presenting cell. A given, albeit fluctuating level of Notch signaling is thus maintained, to preserve the cells in a proliferative, semi-differentiated state, until they encounter the myotube. These observations highlight the unique nature of the myoblasts in their migratory phase. Interestingly, in contrast to the continuous activation of Notch in all swarming myoblasts, a mechanism utilizing the transient nature of interactions between migrating cells has recently been described for the chick somites [123]. Migrating neural crest cells express Delta1 in a mosaic pattern, and while all somitic cells express the receptor, Notch activation is restricted to those which encounter the Delta1-positive neural crest cells passing by. In our analysis, knockdown experiments have shown that Dl is the functional ligand presented by the myoblasts in the context of the pupal myoblast swarm. Mosaic experiments have shown that elevation in the levels of Dl or Ser expressed by the myoblasts, compromise the capacity to receive signals, presumably by cis-inhibition of the receptor. The levels of ligands and receptor expressed by each myoblast thus need to be finely tuned, so that each cell maintains its ability to both send and receive signals. A similar scenario was described during the maturation of the immune synapse. The allogeneic recognition between Th cells and dendritic cells is mediated by bi-directional 77
78 Notch signaling between the cells. In this case, cis-inhibition is prevented by physical segregation of ligands and receptor within the same cell. In one cell type the ligands are at the center and the receptor is at the periphery of the immune synapse, while in the opposing cell they display a complementary distribution [124]. Once the myoblasts are in close proximity to the myotubes, Notch signaling is terminated. This leads to shutdown of Twi, and induction of Sns, D-WIP and most likely a battery of additional genes that are required to execute the fusion process. We assume that the signal(s) to terminate Notch signaling originate from the myotubes, since myoblasts which shut off the Notch reporter and turn on Sns are typically observed in close proximity to the myotubes. The nature of these putative short-range signals terminating Notch signaling is not known. In the satellite cell system, it was suggested that Wnt signaling emanating from the myotube terminates Notch signaling in the transitamplifying cells [125]. Whether this is also true during the formation of the adult flight muscles in Drosophila, remains to be seen. However, regardless of the identity of these molecular mechanisms, considerable similarities in the myogenic programs governing development of the adult fly and vertebrate musculature are clearly evident. 78

79 Figure 8. Notch signaling represses Sns expression and fusion competence in migrating myoblasts. Myoblasts that were set aside during embryogenesis to serve as adult precursors for the indirect flight muscles, are positioned on the wing imaginal disc at the region that will give rise to the notum. During pupariation these cells undergo divisions and migrate as a swarm towards the DLM muscle templates, with which they eventually fuse. The myoblasts are maintained in a precursor state by continuous activation of Notch signaling, triggered by exposure to Dl signals emanating from the adjacent myoblasts in the swarm. Notch activation induces expression of Twi, and suppresses expression of genes essential for fusion such as Sns. We suggest that an unknown signal secreted from the myotube and operating only at close vicinity, leads to termination of Notch signaling in the approaching myoblasts. Induction of Sns expression in a salt and pepper pattern in the myoblasts adjacent to the myotubes, reflects their terminal differentiation and allows them to undergo fusion to the myotube. 79
80 4.4. Methods Reagents list A full list of fly stocks and antibodies used in this work can be found in the supplementary information. Drosophila Genetics UAS-dsRNA constructs were commonly expressed together with UAS-Dicer2, to enhance RNAi activity [87]. Where necessary, the GAL80 ts /TARGET system [126] was used for temporal control of UAS-based transgene expression. Developing flies were maintained at 18 C, allowing for GAL80-based inhibition of GAL4 activity, and shifted at the wandering larvae stage to 29 C, to inactivate GAL80 ts. All Drosophila lines used (mutant alleles, GAL4 drivers and UAS based transgenes) are listed in the Supplementary Material section. Tissue preparation Dissected muscle preparations were obtained from staged pupae (collected as white prepupae at 0 hrs APF). Pupae of the desired age (17-22 hrs APF, unless otherwise specified) were removed from the pupal case, pinned down on Sylgard plates and dissected in cold PBS (see also [1]). Fixation was carried out with 4% para-formaldehyde (PFA) for 30 minutes. Following washes in PBS, the tissue was incubated with antibodies diluted in PBS+0.1% Triton-X and 0.1% bovine serum albumin as blocking reagent. Stained pupal preparations were mounted in 80% glycerol. Primary and secondary antibodies used for immuno-fluorescence are listed in the Supplementary Material section. PLP fixation protocol For staining of Delta, an alternative fixation protocol was utilized. Pupae of the desired age were removed from the pupal case, pinned down on Sylgard plates, dissected in cold PBS and fixed in: 2% PFA, 1.35% lysine and 0.25% Sodium periodate (NaIO 4 ) for 45 minutes. Cell membranes were permeabilized in PBS solution + 0.1% Saponin. All 80
81 washes, primary and secondary antibody incubations were done in PBS + 0.1% Saponin + 5% Normal Goat Serum (NGS) [127]. Clonal Analysis Larvae carrying hs-flp and the act-frt-stop-frt-gal4 cassette were exposed to a mild heat shock in 34ºC for 45 minutes, and allowed to age in 25ºC. Pupae were timed and dissected as mentioned above. Analysis was done on myoblasts that were adjacent to but not fully alighted with the myotubes, to avoid cells that have already begun the fusion process. Whole-mount imaging Pupae of the desired age were removed from the pupal case, and mounted ventrally in 80% glycerol on concave microscope slides. GFP and RFP were imaged immediately after mounting. Immunofluorescence and microscopy Images of immuno-fluorescent samples and live whole mount samples were acquired using Zeiss LSM 710 confocal scanning system using X10 N.A 0.3, X20 N.A 0.5 or X40 N.A 1.1 lenses, and processed using Adobe Photoshop CS3. 81
82 4.5. Supplemental Information to "Fusion competence is repressed in migrating Drosophila myoblasts by Notch signaling" Reagent list Drosophila lines GAL4 drivers: 1151-GAL4 (myoblasts) [128]; rp298 (duf)-gal4 (founder cells) [104]; mef2-gal4 (myogenic mesoderm) [79]; ND637-GAL4 (notum ectoderm) [106]; hs-flp, act-gal-stop-gal, UAS-GFP (flip out clones) [129]. UAS-based transgenes: UAS-myr-mRFP (membrane marker) [130]; UAS-duf [10]; UAS- (TM+C)Duf-FLAG (truncated Duf construct, lacks the extracellular domain) [108]; UAS- CD8GFP (cellular marker) (Bloomington line FB5130); UAS-Delta (Bloomington line FB26694); UAS-Serrate (Bloomington line FB5815); UAS-N ICD (intracellular fragment, constrictively active)[114]; UAS-N ECD (extracellular fragment, dominant negative)[114]; UAS- sns [13]. UAS-dsRNA lines: Wsp (GD13759); Duf/Kirre (GD3111); Rst/IrreC (GD27223); Notch (FB7078); Dl (GD3720); Ser (GD27172); Sns (GD877); Hbs (GD27065 and KK105913). GD and KK lines are from the VDRC (Vienna) collection; FB lines are from Bloomington. Other lines: m6-gfp (Notch activity reporter) [107]; sns-lacz (sns-expression reporter) [110]; rp298(duf)-lacz (expression reporter of duf, expressed in DLMs, weak hypomorphic allele of duf) [72]; hbs-nlacz (expression reporter of hbs) [14]. Antibodies Primary antibodies used included: anti-gfp (chicken, 1:500, Aves), anti-duf (rat, 1:1000, [35]), anti-twist (rabbit, 1:5000, kindly provided by S. Roth); anti-vrp1/d-wip (guinea pig, 1:500, kindly provided by R. H. Palmer [33]); anti-β-galactosidase (mouse, 1:200, Promega); anti-delta (monoclonal mouse, ascites fluid, 1:50, Hybridoma bank C594.9B); anti-serrate (rat, 1:500, kindly provided by K. D. Irvine). Secondary antibodies used included: Alexa Fluor dyes Alexa 405 and Alexa 555 conjugated to anti- mouse, rabbit (Molecular Probes) were used at a dilution of 1:
 at 1:500. Supplementary 1.")
 and GFP expression (green) driven by mef2::gal4. [A,B] Dissected wing discs from third instar larvae.")
83 Anti- mouse, rabbit, guinea-pig, rat conjugated to Cy2, Cy3 and Cy5 (Jackson ImmunoResearch) were diluted 1:400; Atto647N- Phalloidin (Fluka) was used at 2.5µg/ml; anti-chicken conjugated to DyLight 488 (Jackson ImmunoResearch) at 1:500. Supplementary 1. Myoblasts are present in normal numbers on the imaginal wing disc before the onset of migration in duf, rst knockdown larvae. Myoblasts are visualized by Twi expression (red) and GFP expression (green) driven by mef2::gal4. [A,B] Dissected wing discs from third instar larvae. [A] Wild-type myoblasts (mef2-gal4, UAS-CD8GFP,UAS-Dcr2/+) are positioned on the notum of the wing disc. [B] Equivalent numbers of myoblasts are present on the wing disc notum following RNAi-based knockdown of duf and rst (duf rp298/y ; Duf RNAi,rst RNAi /+; mef2-gal4, UAS-CD8GFP,UAS-Dcr2/+). Scale bar- 20µm. Supplementary 2. A dominant-negative truncated form of Duf delays adult myoblast fusion and 83

84 causes abnormal template morphology. Early myoblasts are visualized by Twi expression (green) and fusion-competent myoblasts by D-WIP expression (blue). Myotube templates are marked by Duf (red). [A- H] Dissected pupal muscles hours APF. [A-D] Wild-type myoblasts reach the templates starting at 12 hours APF [A], and fuse to them. As a result, the templates split 16 hours APF [B]. As development progresses, the number of unfused myoblasts decreases [C] and by 24 hours APF almost all myoblasts have fused. [E-H] Expression of a truncated form of the Duf protein (duf:gal4/y; UAS-[TM+C]Duf-FLAG/+) that lacks the extracellular domain, gives rise to a dominant-negative effect on muscle development. [E] Myoblasts migrated normally and reached the myotubes. [F,G] Fusion occurred at a reduced efficiency, manifested in thin myotubes, and accumulation of D-WIP positive, fusion-competent myoblasts outside the myotubes. [H] By the end of adult flight muscle development, some fusion took place, but a significant number of myobalsts still remained unfused. Scale bar for [A-H]- 20µm. Supplementary 3. Notch knockdown inhibits proliferation and expression of Twi in adult myoblasts. [A-D] Dissected pupal muscles 13 and 19 hours APF. Myotubes are marked by Duf expression (blue). [A,B] Wild-type pupae carrying the sns-lacz transgene display a large population of Twi-positive myoblasts (red) and a small population of Sns-positive myoblasts (green), both before [A] and after [B] splitting of the three DLMs. [C,D] Notch knockdown (UAS-N RNAi /+; GAL80 TS /sns-lacz; mef2-gal4,uas- CD8GFP,UAS-Dicer2/+) compromises proliferation and leads to premature differentiation, resulting in a smaller population of myoblasts [C], disappearance of Twi-positive myoblasts by 19 hours APF, and failure of the DLMs to split [D]. Scale bar for [A-D]- 20µm. 84
![Supplementary 4. hibris is positively regulated by Notch signaling, but is not essential for myoblast migration. [A-D] Dissected pupal muscles 18-22 hours APF.](/docs-images/104/165733799/images/85-0.jpg "Myotubes are marked by Duf expression (blue). [A] Ubiquitous expression (red) of the hbs reporter hbs-nlacz(red) in wing disc-derived myoblasts.")
85 Supplementary 4. hibris is positively regulated by Notch signaling, but is not essential for myoblast migration. [A-D] Dissected pupal muscles hours APF. Myotubes are marked by Duf expression (blue). [A] Ubiquitous expression (red) of the hbs reporter hbs-nlacz(red) in wing disc-derived myoblasts. [B] Reducing the levels of Notch signaling by the dominant- negative extracellular domain of Notch (GAL80 TS /hbs-lacz, UAS-N ECD ; mef2-gal4,uas-cd8gfp,uas-dicer2/+) led to a mild reduction in hbs- LacZ expression. [C] Elevation of the levels of Notch signaling by the intracellular domain of Notch (GAL80 TS /hbs-lacz, UAS-N ICD ; mef2-gal4,uas-cd8gfp,uas-dicer2/+) led to a mild increase in hbs- LacZ levels. [D] hbs knockdown by simultaneous expression of two different RNAi constructs (GAL80 TS / UAS-hbs RNAi-KK ; mef2-gal4,uas-cd8gfp,uas-dicer2/uas-hbs RNAi-GD ) did not compromise the ability of myoblasts (marked by Twi- red, and GFP- green) to reach the myotubes (Duf- blue). Fusion to the DLMs occurred in comparable fashion to wild-type (compare to Figure 4A). Scale bars in [A-D] are 20µm. 85
![Supplementary 5. Ectopic expression of Sns does not rescue fusion following constitutive activation of the Notch. [A,B] Dissected pupal muscles 18-22 hours APF.](/docs-images/104/165733799/images/86-0.jpg "Myoblasts are visualized by Twi (red), myotubes by Duf (blue), and GFP (green) marks both cell types.")
86 Supplementary 5. Ectopic expression of Sns does not rescue fusion following constitutive activation of the Notch. [A,B] Dissected pupal muscles hours APF. Myoblasts are visualized by Twi (red), myotubes by Duf (blue), and GFP (green) marks both cell types. [A] Constitutive activation of the Notch pathway (GAL80 TS /UAS-N ICD ; mef2-gal4,uas-cd8gfp,uas-dicer2/+) blocked the myoblast fusion process. [B] Ectopic expression of Sns by the GAL4/UAS system in the background of activated Notch, was not sufficient to rescue the fusion process. Myotubes remained thin and a large number of unfused myoblasts surrounded the muscle templates. It appears therefore that additional genes besides sns, that are essential for fusion, are normally induced following shutdown of Notch signaling. Scale bars for [A-B]- 20µm. 86
אתגר קוביות מחייכות תכולה: 12 קוביות חוברת הוראות ופתרונות ספרון הכולל 60 חידות חידות בדרגות קושי שונות תפיסה חזותית וחשיבה לוגית הקדמה המשחק פרצופים
 אתגר קוביות מחייכות תכולה: 12 קוביות חוברת הוראות ופתרונות ספרון הכולל 60 חידות חידות בדרגות קושי שונות תפיסה חזותית וחשיבה לוגית הקדמה המשחק פרצופים בריבוע מכיל 60 חידות ברמת קושי עולה לשחקן יחיד או שני
אתגר קוביות מחייכות תכולה: 12 קוביות חוברת הוראות ופתרונות ספרון הכולל 60 חידות חידות בדרגות קושי שונות תפיסה חזותית וחשיבה לוגית הקדמה המשחק פרצופים בריבוע מכיל 60 חידות ברמת קושי עולה לשחקן יחיד או שני
שקופית 1
 RESOLUTION Resolution Spatial Resolution Contrast resolution Temporal Resolution Types of Resolution Spatial Resolution also called Detail Resolution the combination of AXIAL and LATERAL resolution -
RESOLUTION Resolution Spatial Resolution Contrast resolution Temporal Resolution Types of Resolution Spatial Resolution also called Detail Resolution the combination of AXIAL and LATERAL resolution -
PowerPoint Presentation
 1 Strangers no more סרט זוכה האוסקר https://www.youtube.com/watch?v=dkciv 4U5Jkw בית הספר היסודי בב"ש http://news.nana10.co.il/article/?articleid= 1017790 שדרת הכניסה לאוניברסיטת תל אביב http://international.tau.ac.il/
1 Strangers no more סרט זוכה האוסקר https://www.youtube.com/watch?v=dkciv 4U5Jkw בית הספר היסודי בב"ש http://news.nana10.co.il/article/?articleid= 1017790 שדרת הכניסה לאוניברסיטת תל אביב http://international.tau.ac.il/
BIG DATA תיאור הקורס המונח Big Data הולך וצובר תאוצה בשנים האחרונות, הוא הופך למגמה רווחת בתעשייה. המשמעות הפרקטית של המונח Big Data הינה טכנולוגיות נ
 BIG DATA תיאור הקורס המונח Big Data הולך וצובר תאוצה בשנים האחרונות, הוא הופך למגמה רווחת בתעשייה. המשמעות הפרקטית של המונח Big Data הינה טכנולוגיות ניתוח וניהול מאגרי מידע בעלי נתונים שאינם מאורגנים,
BIG DATA תיאור הקורס המונח Big Data הולך וצובר תאוצה בשנים האחרונות, הוא הופך למגמה רווחת בתעשייה. המשמעות הפרקטית של המונח Big Data הינה טכנולוגיות ניתוח וניהול מאגרי מידע בעלי נתונים שאינם מאורגנים,
Future Nutrition
 עקומות ב תזונ ה ע תידי ת ד"ר דוד פסיג עתידן, אוניברסיטת בר אילן www.passig.com Futures Studies תחומי חקר ה עתיד - כל כ לות ותעשי ו ת עתיד י ות טכנו לו ג י ו ת עתיד י ות חב ר ות ו ממשל י ם עתיד י ים מדעים
עקומות ב תזונ ה ע תידי ת ד"ר דוד פסיג עתידן, אוניברסיטת בר אילן www.passig.com Futures Studies תחומי חקר ה עתיד - כל כ לות ותעשי ו ת עתיד י ות טכנו לו ג י ו ת עתיד י ות חב ר ות ו ממשל י ם עתיד י ים מדעים
Homework-L9-Skills-1.pub
 My Kriah Homework 1st Grade Level 9: All Skills Much me, energy, and money was invested in developing this program. Therefore reproduc on of this work, whether in it s en rety, in part, or in any form
My Kriah Homework 1st Grade Level 9: All Skills Much me, energy, and money was invested in developing this program. Therefore reproduc on of this work, whether in it s en rety, in part, or in any form
TEL- AVIV UNIVERSITY SACKLER FACULTY OF MEDICINE THE STANLEY STEYER SCHOOL OF HEALTH PROFESSIONS DEPARTMENT OF NURSING אוניברסיטת תל-אביב הפקולטה לרפו
 מס' קורס: 01634949 תכנית המוסמך שיווק במערכות בריאות שיעור, 3 ש"ס מועד הקורס : מרצה: שעות קבלה: סמסטר א', יום ג', 17:00-20:00 דואר אלקטרוני: bcmeir@gmail.com מאיר בן חיים יום ג', 20:00-21:00 )בתאום מראש(
מס' קורס: 01634949 תכנית המוסמך שיווק במערכות בריאות שיעור, 3 ש"ס מועד הקורס : מרצה: שעות קבלה: סמסטר א', יום ג', 17:00-20:00 דואר אלקטרוני: bcmeir@gmail.com מאיר בן חיים יום ג', 20:00-21:00 )בתאום מראש(
Homework Dry 3
 Homework Dry 3 Due date: Sunday, 9/06/2013 12:30 noon Teaching assistant in charge: Anastasia Braginsky Important: this semester the Q&A for the exercise will take place at a public forum only. To register
Homework Dry 3 Due date: Sunday, 9/06/2013 12:30 noon Teaching assistant in charge: Anastasia Braginsky Important: this semester the Q&A for the exercise will take place at a public forum only. To register
Overview of new Office 365 plans for SMBs
 מעבר בין חבילות Online מעבר בין חבילות ב- Open Online to Open Current Plan Upgrade Options Current Plan Upgrade Options Business Essentials Business Premium Enterprise E1 Enterprise E3/E4 Enterprise E1
מעבר בין חבילות Online מעבר בין חבילות ב- Open Online to Open Current Plan Upgrade Options Current Plan Upgrade Options Business Essentials Business Premium Enterprise E1 Enterprise E3/E4 Enterprise E1
Slide 1
 פוליטיקה ארגונית למנהלי משאבי אנוש תמר חושן יעוץ ניהולי "ולחשוב שאני צריכה לבזבז עכשיו כל כך הרבה זמן על הדבר הזה..." למה לי פוליטיקה עכשיו?! "אם אתה מקצוען אמיתי. באמת טוב במה שאתה עושה אתה לא רוצה ולא
פוליטיקה ארגונית למנהלי משאבי אנוש תמר חושן יעוץ ניהולי "ולחשוב שאני צריכה לבזבז עכשיו כל כך הרבה זמן על הדבר הזה..." למה לי פוליטיקה עכשיו?! "אם אתה מקצוען אמיתי. באמת טוב במה שאתה עושה אתה לא רוצה ולא
לקוחות וחברים יקרים,
 מ לקוחות וחברים יקרים, הנדון: סקר בדבר תנאים משפטיים של עסקאות הון סיכון מחצית ראשונה 2022 אנו מתכבדים להציג בפניכם תוצאות הסקר למחצית הראשונה של שנת 0210, המנתח תנאים משפטיים של עסקאות השקעה הון סיכון
מ לקוחות וחברים יקרים, הנדון: סקר בדבר תנאים משפטיים של עסקאות הון סיכון מחצית ראשונה 2022 אנו מתכבדים להציג בפניכם תוצאות הסקר למחצית הראשונה של שנת 0210, המנתח תנאים משפטיים של עסקאות השקעה הון סיכון
טיפול זוגי בגישת AEDP טיפול חווייתי דינמי מואץ Accelarated Experiential Dynamic Psychotherapy
 גישת AEDP - AEDP הטיפול הדינמי-חווייתי-מואץ Accelerated Experiential Dynamic Psychotherapy עקרונות לעשייה טיפולית מצמיחה בטיפול זוגי ופרטני מטפלת זוגית ומשפחתית פעילה במכון הבין-לאומי בגישת AEDP וקיבלה
גישת AEDP - AEDP הטיפול הדינמי-חווייתי-מואץ Accelerated Experiential Dynamic Psychotherapy עקרונות לעשייה טיפולית מצמיחה בטיפול זוגי ופרטני מטפלת זוגית ומשפחתית פעילה במכון הבין-לאומי בגישת AEDP וקיבלה
לקוחות וחברים יקרים, הנדון: סקר בדבר תנאים משפטיים של עסקאות הון סיכון שנת 2022 אנו מתכבדים להציג בפניכם תוצאות הסקר לשנת 0210, המנתח תנאים משפטיים של
 לקוחות וחברים יקרים, הנדון: סקר בדבר תנאים משפטיים של עסקאות הון סיכון שנת 2022 אנו מתכבדים להציג בפניכם תוצאות הסקר לשנת 0210, המנתח תנאים משפטיים של עסקאות השקעה הון סיכון בחברות היי-טק ישראליות ו"ישראליות
לקוחות וחברים יקרים, הנדון: סקר בדבר תנאים משפטיים של עסקאות הון סיכון שנת 2022 אנו מתכבדים להציג בפניכם תוצאות הסקר לשנת 0210, המנתח תנאים משפטיים של עסקאות השקעה הון סיכון בחברות היי-טק ישראליות ו"ישראליות
סדנת תכנות ב C/C++
 פקולטה: מדעי הטבע מחלקה: מדעי המחשב שם הקורס: מבוא למחשבים ושפת C קוד הקורס: 2-7028510 תאריך בחינה: 15.2.2017 משך הבחינה: שעתיים שם המרצה: ד"ר אופיר פלא חומר עזר: פתוח שימוש במחשבון: לא הוראות כלליות:
פקולטה: מדעי הטבע מחלקה: מדעי המחשב שם הקורס: מבוא למחשבים ושפת C קוד הקורס: 2-7028510 תאריך בחינה: 15.2.2017 משך הבחינה: שעתיים שם המרצה: ד"ר אופיר פלא חומר עזר: פתוח שימוש במחשבון: לא הוראות כלליות:
התגוננות בפני כוחות האופל
 ניהול ידע אישי על רגל אחת 1 סוגי ידע Explicit גלוי Tacit סמוי 5 מה אמר השועל לנסיך הקטן? מה קורה בבתי ספר כשמורים או מנהלים עוזבים? 8 ניהול ידע במערכת החינוך רמת התלמיד )ניהול ידע אישי( רמת המורה )ניהול
ניהול ידע אישי על רגל אחת 1 סוגי ידע Explicit גלוי Tacit סמוי 5 מה אמר השועל לנסיך הקטן? מה קורה בבתי ספר כשמורים או מנהלים עוזבים? 8 ניהול ידע במערכת החינוך רמת התלמיד )ניהול ידע אישי( רמת המורה )ניהול
PowerPoint Presentation
 תכנות מתקדם בשפת Java אוניברסיטת תל אביב 1 תוכנה 1 תרגול 3: עבודה עם מחרוזות )Strings( מתודות )Methods( 1 תכנות מתקדם בשפת Java אוניברסיטת תל אביב 2 מחרוזות )STRINGS( 3 מחרוזות String s = Hello ; מחרוזות
תכנות מתקדם בשפת Java אוניברסיטת תל אביב 1 תוכנה 1 תרגול 3: עבודה עם מחרוזות )Strings( מתודות )Methods( 1 תכנות מתקדם בשפת Java אוניברסיטת תל אביב 2 מחרוזות )STRINGS( 3 מחרוזות String s = Hello ; מחרוזות
PowerPoint Presentation
 בינה מלאכותית והמבקר הפנימי לינור דלומי, שותפה, ראש חטיבת ניהול הסיכונים, Deloitte תוכן עניינים מה היא בינה מלאכותית? כשבינה מלאכותית פוגשת את מבקר הפנים האם אתם נותנים למכונה לקבל החלטות עבורכם? מהי
בינה מלאכותית והמבקר הפנימי לינור דלומי, שותפה, ראש חטיבת ניהול הסיכונים, Deloitte תוכן עניינים מה היא בינה מלאכותית? כשבינה מלאכותית פוגשת את מבקר הפנים האם אתם נותנים למכונה לקבל החלטות עבורכם? מהי
מטרות הקורס סמינר מחקר רגשות בשירות סמסטר א' תשע"ט דר' אריק חשין הקורס מיועד להקנות מיומנות בחשיבה מחקרית וביצוע מחקר באמצעו
 מטרות הקורס סמינר מחקר רגשות בשירות סמסטר א' תשע"ט דר' אריק חשין acheshin@univ.haifa.ac.il הקורס מיועד להקנות מיומנות בחשיבה מחקרית וביצוע מחקר באמצעות התנסות בשלבי המחקר השונים. הסמינר יעסוק בנושא רגשות
מטרות הקורס סמינר מחקר רגשות בשירות סמסטר א' תשע"ט דר' אריק חשין acheshin@univ.haifa.ac.il הקורס מיועד להקנות מיומנות בחשיבה מחקרית וביצוע מחקר באמצעות התנסות בשלבי המחקר השונים. הסמינר יעסוק בנושא רגשות
מצגת של PowerPoint
 מבוא כללי לתכנות ולמדעי המחשב תרגול מס' 1 דין שמואל dshmuel110@gmail.com 1 1. מנהלות מרצה: אמיר רובינשטיין, amirr@tau.ac.il שעות קבלה: לשאלות קצרות - מייד לאחר השיעור. ניתן לתאם במייל שעות אחרות. מתרגל:
מבוא כללי לתכנות ולמדעי המחשב תרגול מס' 1 דין שמואל dshmuel110@gmail.com 1 1. מנהלות מרצה: אמיר רובינשטיין, amirr@tau.ac.il שעות קבלה: לשאלות קצרות - מייד לאחר השיעור. ניתן לתאם במייל שעות אחרות. מתרגל:
א.ד.וי.ריידר של עדי ברדה 1. הורדה והתקנה הורד את האפליקציה WD( )AdvRider מהחנות והתקן. למכשירי אנדרואיד בלבד. 2. הגדרות ראשונות לאחר ההתקנה צריך לבצע
AdvRider מהחנות והתקן. למכשירי אנדרואיד בלבד. 2. הגדרות ראשונות לאחר ההתקנה צריך לבצע") א.ד.וי.ריידר של עדי ברדה 1. הורדה והתקנה הורד את האפליקציה WD( )AdvRider מהחנות והתקן. למכשירי אנדרואיד בלבד. 2. הגדרות ראשונות לאחר ההתקנה צריך לבצע באופן חד פעמי, 3 הגדרות פשוטות להתאים את האפליקציה
א.ד.וי.ריידר של עדי ברדה 1. הורדה והתקנה הורד את האפליקציה WD( )AdvRider מהחנות והתקן. למכשירי אנדרואיד בלבד. 2. הגדרות ראשונות לאחר ההתקנה צריך לבצע באופן חד פעמי, 3 הגדרות פשוטות להתאים את האפליקציה
HUJI Syllabus
 Syllabus INTRODUCTION TO SOCIAL WORK-II - 3103 Last update 30-10-2016 HU Credits: 2 Degree/Cycle: 1st degree (Bachelor) Responsible Department: social work Academic year: 0 Semester: 2nd Semester Teaching
Syllabus INTRODUCTION TO SOCIAL WORK-II - 3103 Last update 30-10-2016 HU Credits: 2 Degree/Cycle: 1st degree (Bachelor) Responsible Department: social work Academic year: 0 Semester: 2nd Semester Teaching
Why to contact <NORAIL.EMEKREFAIM.GMAIL.COM>
 PART OF EHUD OLMERT S LEGACY A ROLE MODEL FOR TODAY Mazel pushed some lightstands into the pool, causing a short-circuit and disabling the light, and then told the artist that: "This is not a work of art!
PART OF EHUD OLMERT S LEGACY A ROLE MODEL FOR TODAY Mazel pushed some lightstands into the pool, causing a short-circuit and disabling the light, and then told the artist that: "This is not a work of art!
w-2003
 Draft SI 1003 part 1.1 January 2013 טיוטה לתקן ישראלי ת"י 1003 חלק 1.1 ינואר 2013 ICS CODE: שחרור עופרת וקדמיום מכלים הבאים במגע עם מזון: כלים קרמיים, כלים קרמייםזכוכיתיים וכלי הגשה מזכוכית שיטת בדיקה
Draft SI 1003 part 1.1 January 2013 טיוטה לתקן ישראלי ת"י 1003 חלק 1.1 ינואר 2013 ICS CODE: שחרור עופרת וקדמיום מכלים הבאים במגע עם מזון: כלים קרמיים, כלים קרמייםזכוכיתיים וכלי הגשה מזכוכית שיטת בדיקה
eriktology The Prophets Book of 1 st Kings [1]
![eriktology The Prophets Book of 1 st Kings [1]](/thumbs/99/139389989.jpg "eriktology The Prophets Book of 1 st Kings [1]") eriktology The Prophets Book of 1 st Kings [1] [2] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement
eriktology The Prophets Book of 1 st Kings [1] [2] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement
מקביליות
 PROMELA גרא וייס המחלקה למדעי המחשב אוניברסיטת בן-גוריון עדכון אחרון: 21:40 15/06/2013 2 שפת מ פ ר ט עם ס מ נ ט יק ה מוגדרת באופן מתמטי "שפת תכנות" למודלים המטרה: לאפשר גם לכאלה שאינם חוקרים בתחום לבנות
PROMELA גרא וייס המחלקה למדעי המחשב אוניברסיטת בן-גוריון עדכון אחרון: 21:40 15/06/2013 2 שפת מ פ ר ט עם ס מ נ ט יק ה מוגדרת באופן מתמטי "שפת תכנות" למודלים המטרה: לאפשר גם לכאלה שאינם חוקרים בתחום לבנות
שקופית 1
 www.pwc.com/il חידושים בתחום התמריצים טלי ברנד, רו"ח, דירקטורית, מנהלת מחלקת תמריצים, אוקטובר 2014 תוכן העניינים דגשים לגבי הטבות מס על פי החוק לעידוד השקעות הון תכניות מענקי מחקר ופיתוח נבחרות שינויים
www.pwc.com/il חידושים בתחום התמריצים טלי ברנד, רו"ח, דירקטורית, מנהלת מחלקת תמריצים, אוקטובר 2014 תוכן העניינים דגשים לגבי הטבות מס על פי החוק לעידוד השקעות הון תכניות מענקי מחקר ופיתוח נבחרות שינויים
Slide 1
 1 אובייקטים היום בתרגול: 2.)objects מחלקות )classes( ואובייקטים )מופעים, )fields( שדות המחלקה שיטות הכמסה )methods של מחלקה. ( class מחלקה - עד עכשיו השתמשנו בעיקר בטיפוסים מובנים ופונקציות המבצעות חישובים
1 אובייקטים היום בתרגול: 2.)objects מחלקות )classes( ואובייקטים )מופעים, )fields( שדות המחלקה שיטות הכמסה )methods של מחלקה. ( class מחלקה - עד עכשיו השתמשנו בעיקר בטיפוסים מובנים ופונקציות המבצעות חישובים
<4D F736F F D20EEE5F4FA20FAF7F6E9F820E4EEF9F4E7E420F9ECE420E4EEF9F4E7E420F9ECE5>
 המשפחה שלה, המשפחה שלו - עמדות מתבגרות ומתבגרים ביחס למעמד האשה במשפחה, כפונקציה של משפחת המוצא והיבטים מעודדי סובלנות של ההקשר החינוכי בלהה אדמנית, מכללת 'תלפיות' במחקר זה נבחנו עמדותיהם של מתבגרות ומתבגרים
המשפחה שלה, המשפחה שלו - עמדות מתבגרות ומתבגרים ביחס למעמד האשה במשפחה, כפונקציה של משפחת המוצא והיבטים מעודדי סובלנות של ההקשר החינוכי בלהה אדמנית, מכללת 'תלפיות' במחקר זה נבחנו עמדותיהם של מתבגרות ומתבגרים
Microsoft PowerPoint - CE_Candidates_2011.ppt [Compatibility Mode]
![Microsoft PowerPoint - CE_Candidates_2011.ppt [Compatibility Mode]](/thumbs/101/150419256.jpg "Microsoft PowerPoint - CE_Candidates_2011.ppt [Compatibility Mode]") תשע"בב פתוח ו וירטואלי לקראת שנת הלמוד הלימודים יום ראשון ב- תואר מחשבים הנדסת הלימודים שנת לקראת הוירטואלי הפתוח ליום הבאים ברוכים מחשבים הנדסת עלל מקצוע פרטים מספר זוו תמצאו תשס"בב. במצגת וי שיש שינויים
תשע"בב פתוח ו וירטואלי לקראת שנת הלמוד הלימודים יום ראשון ב- תואר מחשבים הנדסת הלימודים שנת לקראת הוירטואלי הפתוח ליום הבאים ברוכים מחשבים הנדסת עלל מקצוע פרטים מספר זוו תמצאו תשס"בב. במצגת וי שיש שינויים
ניטול ידני
 הכנס ה- 17 של המוסד לבטיחות ולגהות 17 נובמבר 2014 בטיחות ובריאות בתעסוקה הרמה וניטול ידני: ד"ר יוהנה גייגר ארגונומית ארצית המוסד לבטיחות ולגהות ענבר גלבוע ארגונומית מחוזית המוסד לבטיחות ולגהות ניטול ידני
הכנס ה- 17 של המוסד לבטיחות ולגהות 17 נובמבר 2014 בטיחות ובריאות בתעסוקה הרמה וניטול ידני: ד"ר יוהנה גייגר ארגונומית ארצית המוסד לבטיחות ולגהות ענבר גלבוע ארגונומית מחוזית המוסד לבטיחות ולגהות ניטול ידני
Microsoft PowerPoint - 59.pptx
 Anterior Knee Pain + Syndrome מקרה שהיה... מאת: יצחק בזזינסקי סיני הצגת מקרים רפואת ספורט שנה ג בן 32, עובד כמנהל שיווק בעבודה משרדית עם הרבה הליכות ומדרגות בריא בדר"כ עוסק בספורט חובבני- כדורגל וריצות
Anterior Knee Pain + Syndrome מקרה שהיה... מאת: יצחק בזזינסקי סיני הצגת מקרים רפואת ספורט שנה ג בן 32, עובד כמנהל שיווק בעבודה משרדית עם הרבה הליכות ומדרגות בריא בדר"כ עוסק בספורט חובבני- כדורגל וריצות
תרגול מס' 1
 תרגול 6 הסתעפויות 1 מבוסס על שקפים מאת יאן ציטרין קפיצות לא מותנות Unconditional Branch br label PC לאחר ה- fetch של פקודת ה- branch PC לאחר הביצוע של פקודת ה- branch pc label br label הקפיצה מתבצעת תמיד,
תרגול 6 הסתעפויות 1 מבוסס על שקפים מאת יאן ציטרין קפיצות לא מותנות Unconditional Branch br label PC לאחר ה- fetch של פקודת ה- branch PC לאחר הביצוע של פקודת ה- branch pc label br label הקפיצה מתבצעת תמיד,
eriktology The Writings Book of Proverbs [1]
![eriktology The Writings Book of Proverbs [1]](/thumbs/99/141036036.jpg "eriktology The Writings Book of Proverbs [1]") eriktology The Writings Book of Proverbs [1] [2] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement that
eriktology The Writings Book of Proverbs [1] [2] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement that
eriktology The Writings Book of Psalms [1]
![eriktology The Writings Book of Psalms [1]](/thumbs/99/139819849.jpg "eriktology The Writings Book of Psalms [1]") eriktology The Writings Book of Psalms [1] [2] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement that
eriktology The Writings Book of Psalms [1] [2] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement that
eriktology The Prophets Book of Isaiah [1]
![eriktology The Prophets Book of Isaiah [1]](/thumbs/98/138126750.jpg "eriktology The Prophets Book of Isaiah [1]") eriktology The Prophets Book of Isaiah [1] [2] [3] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement
eriktology The Prophets Book of Isaiah [1] [2] [3] FOREWORD It should be noted when using this workbook, that we ( Eric, Lee, James, and a host of enthusiastic encouragers ) are not making a statement
ארסמוס+ עדכון
 הדגשים לקול הקורא 2018 Capacity Building Capacity Building "בניית יכולות של מערכת ההשכלה הגבוהה הישראלית תוך שיתוף פעולה בינלאומי" מוסדות להשכלה גבוהה פיתוח תכניות לימוד בגוון דיסציפלינות קידום בינלאומיות
הדגשים לקול הקורא 2018 Capacity Building Capacity Building "בניית יכולות של מערכת ההשכלה הגבוהה הישראלית תוך שיתוף פעולה בינלאומי" מוסדות להשכלה גבוהה פיתוח תכניות לימוד בגוון דיסציפלינות קידום בינלאומיות
מסמך1
 Draft of SI 9612 June 2019 טיוטה לתקן הישראלי ת"י 9612 ISO 9612 - Second edition: 2009-04-01 יוני 2019 ICS CODE: 13.140 אקוסטיקה: קביעה של רמת חשיפה לרעש בסביבת עבודה שיטה הנדסית Acoustics Determination
Draft of SI 9612 June 2019 טיוטה לתקן הישראלי ת"י 9612 ISO 9612 - Second edition: 2009-04-01 יוני 2019 ICS CODE: 13.140 אקוסטיקה: קביעה של רמת חשיפה לרעש בסביבת עבודה שיטה הנדסית Acoustics Determination
<4D F736F F F696E74202D EEE420E6E420E5ECEEE920E6E4202D20F4F8E5F420F1E9EEE5EF>
 אבחון טרום ההשרשה אלכס סימון פרופ' מחלקת נשים ויולדות היחידה להפריה חו ץ גופית בית החולים הדסה חלק מהתמונות והאיורים במצגת זו נלקחו מספרים ואתרים באינטרנט והשימוש בהם נעשה למטרות לימוד בלבד. כל הזכויות
אבחון טרום ההשרשה אלכס סימון פרופ' מחלקת נשים ויולדות היחידה להפריה חו ץ גופית בית החולים הדסה חלק מהתמונות והאיורים במצגת זו נלקחו מספרים ואתרים באינטרנט והשימוש בהם נעשה למטרות לימוד בלבד. כל הזכויות
אוניברסיטת בר אילן - הפקולטה לרפואה בגליל
 אוניברסיטת בר אילן - הפקולטה לרפואה בגליל "הקשר בין DAT פולימורפיזם ועיבוד רגשות מחקר ראשוני" בסכיזופרניה. "Association between DAT polymorphism and mentalization in schizophrenia; a pilot study" עבודת
אוניברסיטת בר אילן - הפקולטה לרפואה בגליל "הקשר בין DAT פולימורפיזם ועיבוד רגשות מחקר ראשוני" בסכיזופרניה. "Association between DAT polymorphism and mentalization in schizophrenia; a pilot study" עבודת
הליבה של פסיכיאטריה ונחיצות הפסיכיאטריה בחברה חופשית ומתוקנת
 הליבה של הפסיכיאטריה ונחיצות הפסיכיאטריה בחברה חופשית ומתוקנת פרופ' חיים בלמקר יו"ר איגוד הפסיכיאטריה בישראל בעל טור בעיתון הארץ קבע ששנת 2016 היתה הכי קשה לצה"ל מבחינת התקשורת מאז הקמתו כנראה שהפסיכיאטריה
הליבה של הפסיכיאטריה ונחיצות הפסיכיאטריה בחברה חופשית ומתוקנת פרופ' חיים בלמקר יו"ר איגוד הפסיכיאטריה בישראל בעל טור בעיתון הארץ קבע ששנת 2016 היתה הכי קשה לצה"ל מבחינת התקשורת מאז הקמתו כנראה שהפסיכיאטריה
פתרון מוצע לבחינת מה"ט ב_שפת c מועד ב אביב תשע"ט, אפריל 2019 מחברת: גב' זהבה לביא, מכללת אורט רחובות שאלה מספר 1 מוגדרת מחרוזת המורכבת מהספרות 0 עד 9.
 פתרון מוצע לבחינת מה"ט ב_שפת c מועד ב אביב תשע"ט, אפריל 2019 מחברת: גב' זהבה לביא, מכללת אורט רחובות שאלה מספר 1 מוגדרת מחרוזת המורכבת מהספרות 0 עד 9. הדפסה ראשונה: מתבצעת לולאה שרצה מאפס עד אורך המחרוזת.
פתרון מוצע לבחינת מה"ט ב_שפת c מועד ב אביב תשע"ט, אפריל 2019 מחברת: גב' זהבה לביא, מכללת אורט רחובות שאלה מספר 1 מוגדרת מחרוזת המורכבת מהספרות 0 עד 9. הדפסה ראשונה: מתבצעת לולאה שרצה מאפס עד אורך המחרוזת.
_v18A
 Draft SI 1003 part 1.2 January 2013 טיוטה לתקן ישראלי ת"י 1003 חלק 1.2 ינואר 2013 ICS CODE: שחרור עופרת וקדמיום מכלים הבאים במגע עם מזון: כלים קרמיים, כלים קרמיים-זכוכיתיים וכלי הגשה מזכוכית גבולות מותרים
Draft SI 1003 part 1.2 January 2013 טיוטה לתקן ישראלי ת"י 1003 חלק 1.2 ינואר 2013 ICS CODE: שחרור עופרת וקדמיום מכלים הבאים במגע עם מזון: כלים קרמיים, כלים קרמיים-זכוכיתיים וכלי הגשה מזכוכית גבולות מותרים
_20
 Draft SI 17022 August 2019 טיוטה לתקן ישראלי ת"י 17022 אוגוסט 2019 ICS CODE: 97.190 מוצרים לטיפול בילדים עזרים לרחצה ושיטות בדיקה דרישות בטיחות Child care articles - Bathing aids - Safety requirements
Draft SI 17022 August 2019 טיוטה לתקן ישראלי ת"י 17022 אוגוסט 2019 ICS CODE: 97.190 מוצרים לטיפול בילדים עזרים לרחצה ושיטות בדיקה דרישות בטיחות Child care articles - Bathing aids - Safety requirements
מהוא לתכנות ב- JAVA מעבדה 3
 מבוא לתכנות ב- JAVA מעבדה 3 נושאי התרגול לולאות ניפוי שגיאות לולאות - הקדמה כיצד הייתם כותבים תוכנית שתדפיס את המספרים השלמים בין 1 ל- 100 בעזרת הכלים שלמדתם עד עתה? חייבת להיות דרך אחרת מאשר לכתוב 100
מבוא לתכנות ב- JAVA מעבדה 3 נושאי התרגול לולאות ניפוי שגיאות לולאות - הקדמה כיצד הייתם כותבים תוכנית שתדפיס את המספרים השלמים בין 1 ל- 100 בעזרת הכלים שלמדתם עד עתה? חייבת להיות דרך אחרת מאשר לכתוב 100
תרגיל 3 בקורס תוכנה 1
 1 תוכנה תרגיל מספר 8 הנחיות כלליות: קראו בעיון את קובץ נוהלי הגשת התרגילים אשר נמצא באתר הקורס. הגשת התרגיל תעשה במערכת ה VirtualTAU בלבד.(http://virtual2002.tau.ac.il/) יש להגיש קובץ zip יחיד הנושא את
1 תוכנה תרגיל מספר 8 הנחיות כלליות: קראו בעיון את קובץ נוהלי הגשת התרגילים אשר נמצא באתר הקורס. הגשת התרגיל תעשה במערכת ה VirtualTAU בלבד.(http://virtual2002.tau.ac.il/) יש להגיש קובץ zip יחיד הנושא את
מבוא למדעי המחשב
 מבוא למדעי המחשב מחרוזות, חתימה של פונקציה ומעטפות תוכן עניינים טיפוסים מורכבים טיפוסים מורכבים ערך שם טיפוס 12 m int undef. x boolean true y boolean arr int[] כאלה שעשויים להכיל יותר מערך פרימיטיבי אחד
מבוא למדעי המחשב מחרוזות, חתימה של פונקציה ומעטפות תוכן עניינים טיפוסים מורכבים טיפוסים מורכבים ערך שם טיפוס 12 m int undef. x boolean true y boolean arr int[] כאלה שעשויים להכיל יותר מערך פרימיטיבי אחד
פרקים נבחרים בהדברה ביולוגית ומשולבת בתות-שדה
 שימוש בפרוקי רגליים מועילים בחקלאות המודרנית: מציאות, סיכויים ותהיות שמעון שטיינברג, ביו-בי שדה אליהו בע"מ הדברה משולבת. Integrated Pest Management הגדרה: ממשק המנצל לאורך זמן את כל הטכניקות המתאימות,
שימוש בפרוקי רגליים מועילים בחקלאות המודרנית: מציאות, סיכויים ותהיות שמעון שטיינברג, ביו-בי שדה אליהו בע"מ הדברה משולבת. Integrated Pest Management הגדרה: ממשק המנצל לאורך זמן את כל הטכניקות המתאימות,
(Microsoft PowerPoint \372\347\370\345\372 \341\362\351\343\357 \344\343\351\342\351\350\354\351)
") אוניברסיטת תל-אביב הפקולטה לניהול ע"ש קולר מכון אלי הורביץ לניהול אסטרטגי ברוכים הבאים לכנס מקצועי בנושא: 14 ליוני, 2018 אוניברסיטת תל - אביב הפקולטה לניהול ע"ש קולר מכון אלי הורביץ לניהול אסטרטגי מנהלי
אוניברסיטת תל-אביב הפקולטה לניהול ע"ש קולר מכון אלי הורביץ לניהול אסטרטגי ברוכים הבאים לכנס מקצועי בנושא: 14 ליוני, 2018 אוניברסיטת תל - אביב הפקולטה לניהול ע"ש קולר מכון אלי הורביץ לניהול אסטרטגי מנהלי
GO! חוקי המשחק 2-5 שחקנים גילאי 8+ מיוצר בסין עבור פוקסמיינד. משווק בישראל על ידי פוקסמיינד בע"מ. כפר ורדים, מרווה 12 ח.פ טל Mad
 GO! חוקי המשחק 2-5 שחקנים גילאי 8+ מיוצר בסין עבור פוקסמיינד. משווק בישראל על ידי פוקסמיינד בע"מ. כפר ורדים, מרווה 12 ח.פ. 513226795 טל 03-9092034 Made in China for FoxMind. Distributed in Israel by FoxMind
GO! חוקי המשחק 2-5 שחקנים גילאי 8+ מיוצר בסין עבור פוקסמיינד. משווק בישראל על ידי פוקסמיינד בע"מ. כפר ורדים, מרווה 12 ח.פ. 513226795 טל 03-9092034 Made in China for FoxMind. Distributed in Israel by FoxMind
ח ס פ - י ש י ז ר א מ w w w. p t o r a. c o. i l S a l e p t o r a. c o. i l
 2 0 1 9 ח ס פ - י ש י ז ר א מ 054-9852860 w w w p t o r a c o i l S a l e s @ p t o r a c o i l Sack Of Nuts Haroset in a box מ א ר ז מ ת ו ק ו י י ח ו ד י : צ נ צ נ ת ח ר ו ס ת מ ד ב ר י ת מ י ו ח ד ת
2 0 1 9 ח ס פ - י ש י ז ר א מ 054-9852860 w w w p t o r a c o i l S a l e s @ p t o r a c o i l Sack Of Nuts Haroset in a box מ א ר ז מ ת ו ק ו י י ח ו ד י : צ נ צ נ ת ח ר ו ס ת מ ד ב ר י ת מ י ו ח ד ת
PowerPoint Presentation
 תוכנה 1 3 תרגול מס' מתודות ותיכון לפי חוזים חוזה בין ספק ללקוח חוזה בין ספק ללקוח מגדיר עבור כל שרות: תנאי ללקוח - "תנאי קדם" precondition - - תנאי לספק "תנאי אחר".postcondition לקוח ספק 2 תנאי קדם )preconditions(
תוכנה 1 3 תרגול מס' מתודות ותיכון לפי חוזים חוזה בין ספק ללקוח חוזה בין ספק ללקוח מגדיר עבור כל שרות: תנאי ללקוח - "תנאי קדם" precondition - - תנאי לספק "תנאי אחר".postcondition לקוח ספק 2 תנאי קדם )preconditions(
מבוא למדעי המחשב
 מבוא למדעי המחשב הממשקים Iterable,Set ו- Iterator תוכן עניינים מבנה נתונים מבנה נתונים מבנה נתונים הוא container של עצמים שמוגדרות עליו מספר פעולות למשל קבוצה של עצמים (אוסף ללא חזרות) עם הפעולות: הוספת
מבוא למדעי המחשב הממשקים Iterable,Set ו- Iterator תוכן עניינים מבנה נתונים מבנה נתונים מבנה נתונים הוא container של עצמים שמוגדרות עליו מספר פעולות למשל קבוצה של עצמים (אוסף ללא חזרות) עם הפעולות: הוספת
<4D F736F F D20E1F2E9E5FA20F2E9F7F8E9E5FA20E1E2E9EC20E4F8EA2D20F1E9ECE1E5F120E5E1E9E1ECE9E5E2F8F4E9E F484C>
 אוניברסיטת בן-גוריון בנגב המחלקה לפסיכולוגיה בעיות עיקריות בגיל הרך: האכלה, שינה והרגלי ניקיון 2017 דר' נעמה עצבה-פוריה שעת קבלה: יום חמישי 8-9 (בתיאום מראש) חדר: 214 בניין 98 מתרגלת: עטרה מנשה (menashe@bgu.ac.il)
אוניברסיטת בן-גוריון בנגב המחלקה לפסיכולוגיה בעיות עיקריות בגיל הרך: האכלה, שינה והרגלי ניקיון 2017 דר' נעמה עצבה-פוריה שעת קבלה: יום חמישי 8-9 (בתיאום מראש) חדר: 214 בניין 98 מתרגלת: עטרה מנשה (menashe@bgu.ac.il)
נגיפים ד"ר שונית גלזר
 נגיפים ד"ר שונית גלזר ניתן למיין מיקרואורגניזמים לא תאיים cellular Non תאיים - Cellular אאוקריוטים - Eukaryotes פרוקריוטים - prokaryotes נגיפים Viruses( ) פטריות חיידקים וירואידים טפילים ארכיאה פריונים
נגיפים ד"ר שונית גלזר ניתן למיין מיקרואורגניזמים לא תאיים cellular Non תאיים - Cellular אאוקריוטים - Eukaryotes פרוקריוטים - prokaryotes נגיפים Viruses( ) פטריות חיידקים וירואידים טפילים ארכיאה פריונים
Counting the Omer Leviticus 23:15-16 And from the day on which you bring the sheaf of elevation offering the day after the Sabbath you shall count off
 Counting the Omer Leviticus 23:15-16 And from the day on which you bring the sheaf of elevation offering the day after the Sabbath you shall count off seven weeks. They must be complete: you must count
Counting the Omer Leviticus 23:15-16 And from the day on which you bring the sheaf of elevation offering the day after the Sabbath you shall count off seven weeks. They must be complete: you must count
תרגול 1
 מבוא למדעי המחשב 2019 תרגול 5 מחרוזות, חתימות ורקורסיה מחרוזות רצף של תווים רקורסיה קריאה של מתודה לעצמה באופן ישיר או עקיף ראינו בהרצאה מחרוזות: תווים, חתימות: העמסה- String,הצהרה, overloading אתחול רקורסיה:
מבוא למדעי המחשב 2019 תרגול 5 מחרוזות, חתימות ורקורסיה מחרוזות רצף של תווים רקורסיה קריאה של מתודה לעצמה באופן ישיר או עקיף ראינו בהרצאה מחרוזות: תווים, חתימות: העמסה- String,הצהרה, overloading אתחול רקורסיה:
פיסיקה 1 ב' מרצים: גולן בל, משה שכטר, מיכאל גדלין מועד ב משך המבחן 3 שעות חומר עזר: דף נוסחאות מצורף, מחשבון אסור בהצלחה! חלק א'
 פיסיקה 1 ב' 203-1-1391 מרצים: גולן בל, משה שכטר, מיכאל גדלין מועד ב 03.08.2017 משך המבחן 3 שעות חומר עזר: דף נוסחאות מצורף, מחשבון אסור בהצלחה! חלק א' - שאלות אמריקאיות (כל שאלה - 5 נק') - יש לסמן תשובה
פיסיקה 1 ב' 203-1-1391 מרצים: גולן בל, משה שכטר, מיכאל גדלין מועד ב 03.08.2017 משך המבחן 3 שעות חומר עזר: דף נוסחאות מצורף, מחשבון אסור בהצלחה! חלק א' - שאלות אמריקאיות (כל שאלה - 5 נק') - יש לסמן תשובה
שקופית 1
 הזהות הדיגיטלית ומות הפרטיות דוגמא מתחום התקשורת נעם למלשטריך לטר דיקאן מיסד בית הספר לתקשורת הבינתחומי הרצליה כנס העצמה דיגיטלית : מושגים ישנים-משמעויות חדשות 3.7.12 1 תקן בינלאומי לזהויות דיגיטליות קיים
הזהות הדיגיטלית ומות הפרטיות דוגמא מתחום התקשורת נעם למלשטריך לטר דיקאן מיסד בית הספר לתקשורת הבינתחומי הרצליה כנס העצמה דיגיטלית : מושגים ישנים-משמעויות חדשות 3.7.12 1 תקן בינלאומי לזהויות דיגיטליות קיים
רשתות חברתיות לאקדמיה- ידע ומידע מדעי וחלון הזדמנויות לספריות
 רשתות חברתיות לאקדמיה- ידע ומידע מדעי וחלון הזדמנויות לספריות דר' יפה אהרוני רשת חברתית הגדרה- webopedia Abbreviated as SNS a social networking site is the phrase used to describe any Web site that enables
רשתות חברתיות לאקדמיה- ידע ומידע מדעי וחלון הזדמנויות לספריות דר' יפה אהרוני רשת חברתית הגדרה- webopedia Abbreviated as SNS a social networking site is the phrase used to describe any Web site that enables
דוגמאות שהוצגו בהרצאה 10 בקורס יסודות מערכות פתוחות דוגמה 1 דוגמאות של פונקציות ב- awk שמראות שהעברת פרמטרים של משתנים פשוטים היא by value והעברת פרמט
 דוגמאות שהוצגו בהרצאה 10 בקורס יסודות מערכות פתוחות דוגמה 1 דוגמאות של פונקציות ב- awk שמראות שהעברת פרמטרים של משתנים פשוטים היא by value והעברת פרמטרים של מערכים היא by reference וכן דוגמאות שמראות שמשתנים
דוגמאות שהוצגו בהרצאה 10 בקורס יסודות מערכות פתוחות דוגמה 1 דוגמאות של פונקציות ב- awk שמראות שהעברת פרמטרים של משתנים פשוטים היא by value והעברת פרמטרים של מערכים היא by reference וכן דוגמאות שמראות שמשתנים
אוניברסיטת בן-גוריון בנגב NEGEV THE BEN-GURION UNIVERSITY OF
 אוניברסיטת בן-גוריון בנגב הפקולטה למדעי הרוח והחברה טופס סילבוס לסטודנט תשע"ז 2016- המחלקה ל - חינוך סמסטר א' שם הקורס: התמודדות עם בעיות התנהגות במסגרות החינוך- באוריינטציה מערכתית. 12920653 מס' קורס
אוניברסיטת בן-גוריון בנגב הפקולטה למדעי הרוח והחברה טופס סילבוס לסטודנט תשע"ז 2016- המחלקה ל - חינוך סמסטר א' שם הקורס: התמודדות עם בעיות התנהגות במסגרות החינוך- באוריינטציה מערכתית. 12920653 מס' קורס
<4D F736F F D20FAE5EBF0E9FA20EEE0F1E8F8>
 תוכנית הכנס המדעי *כפופה לשינויים ועדה מדעית: ד"ר איריס מנור, פרופ' גיל זלצמן, ד"ר איתי ברגר, ד"ר דיצה צחור, ד"ר רחל בן חיון, ד"ר שמואל הירשמן, פרופ' יהודית אהרון-פרץ ועדה מארגנת: ד"ר מרים פסקין, פרופ'
תוכנית הכנס המדעי *כפופה לשינויים ועדה מדעית: ד"ר איריס מנור, פרופ' גיל זלצמן, ד"ר איתי ברגר, ד"ר דיצה צחור, ד"ר רחל בן חיון, ד"ר שמואל הירשמן, פרופ' יהודית אהרון-פרץ ועדה מארגנת: ד"ר מרים פסקין, פרופ'
תהליך הגשה והנחיות כתיבה לעבודת גמר / תזה פרקים הקדמה תהליך הגשת עבודת המחקר ואישורה הנחיות תוכן לעבודת המחקר הנחיות כתיבה לעבודת המחקר הקדמה במסגרת ל
 תהליך הגשה והנחיות כתיבה לעבודת גמר / תזה פרקים הקדמה תהליך הגשת עבודת המחקר ואישורה הנחיות תוכן לעבודת המחקר הנחיות כתיבה לעבודת המחקר הקדמה במסגרת לימודי החוג לפסיכולוגיה תואר שני במרכז האוניברסיטאי
תהליך הגשה והנחיות כתיבה לעבודת גמר / תזה פרקים הקדמה תהליך הגשת עבודת המחקר ואישורה הנחיות תוכן לעבודת המחקר הנחיות כתיבה לעבודת המחקר הקדמה במסגרת לימודי החוג לפסיכולוגיה תואר שני במרכז האוניברסיטאי
2
 אוניברסיטת תל-אביב הפקולטה למדעי הרוח ע"ש לסטר וסאלי אנטין בית הספר למדעי התרבות ע"ש שירלי ולסלי פורטר היחידה למחקר התרבות הנגרות ביישוב היהודי בארץ-ישראל 9191-9191: מקרה לבחינת התארגנות של שדה פעולה מקצועי
אוניברסיטת תל-אביב הפקולטה למדעי הרוח ע"ש לסטר וסאלי אנטין בית הספר למדעי התרבות ע"ש שירלי ולסלי פורטר היחידה למחקר התרבות הנגרות ביישוב היהודי בארץ-ישראל 9191-9191: מקרה לבחינת התארגנות של שדה פעולה מקצועי
95% בוגרים להשכלה גבוהה 70% דור ראשון במשפחתם יוזמה להקמת בית ספר תיכון על פי מודל ה - Picture Big תמונההגדולהה
 95% בוגרים להשכלה גבוהה 70% דור ראשון במשפחתם יוזמה להקמת בית ספר תיכון על פי מודל ה - Picture Big תמונההגדולהה על מה אובמה מדבר? מודל תומך למידה ה- Met הוא שמו של בית ספר תיכון ברוד-איילנד, ארה ב, שמשמש
95% בוגרים להשכלה גבוהה 70% דור ראשון במשפחתם יוזמה להקמת בית ספר תיכון על פי מודל ה - Picture Big תמונההגדולהה על מה אובמה מדבר? מודל תומך למידה ה- Met הוא שמו של בית ספר תיכון ברוד-איילנד, ארה ב, שמשמש
Guide For Examinees Inter-University Psychometric Entrance Test HEBREW PROFICIENCY TEST (YAEL) This test is intended for examinees who take the Psycho
 This test is intended for examinees who take the Psycho") Guide For Examinees Inter-University Psychometric Entrance Test HEBREW PROFICIENCY TEST (YAEL) This test is intended for examinees who take the Psychometric Entrance Test in a language other than Hebrew.
Guide For Examinees Inter-University Psychometric Entrance Test HEBREW PROFICIENCY TEST (YAEL) This test is intended for examinees who take the Psychometric Entrance Test in a language other than Hebrew.
LTEY leaflet
 Index 2 3 5 7 Letter from Rav Noam Rationale and Goals Global Activities Ideas for Programs The night of Torat Eretz Yisrael 1 בעזרתו יתברך ובהשתדלותנו יז שב"ט תשע"ג שליחינו סביב הגלובוס שלומות! לאחר הפסגות
Index 2 3 5 7 Letter from Rav Noam Rationale and Goals Global Activities Ideas for Programs The night of Torat Eretz Yisrael 1 בעזרתו יתברך ובהשתדלותנו יז שב"ט תשע"ג שליחינו סביב הגלובוס שלומות! לאחר הפסגות
מבוא לתכנות ב- JAVA תרגול 11
 מבוא לתכנות ב- JAVA תרגול 11 רשימה מקושרת אוסף סדור של איברים מאותו טיפוס. קודקוד ברשימה )Node( מכיל את המידע + הצבעה לקודקוד הבא ברשימה data next first רשימה :)List( מיוצגת ע"י מצביע לאיבר הראשון ברשימה
מבוא לתכנות ב- JAVA תרגול 11 רשימה מקושרת אוסף סדור של איברים מאותו טיפוס. קודקוד ברשימה )Node( מכיל את המידע + הצבעה לקודקוד הבא ברשימה data next first רשימה :)List( מיוצגת ע"י מצביע לאיבר הראשון ברשימה
תורת הקומפילציה
 תורת הקומפילציה תרגיל בית 2 הוראות לתרגיל 2 בשפת MINI-PASCAL הפרוייקט המצורף הינו קוד שלד של מהדר לשפת mini-pascal עליכם לממש בסביבת (Eclipse (Java את הפונקציות המתאימות לפי החומר שנלמד בהרצאה ע"מ שהמהדר
תורת הקומפילציה תרגיל בית 2 הוראות לתרגיל 2 בשפת MINI-PASCAL הפרוייקט המצורף הינו קוד שלד של מהדר לשפת mini-pascal עליכם לממש בסביבת (Eclipse (Java את הפונקציות המתאימות לפי החומר שנלמד בהרצאה ע"מ שהמהדר
הגשה תוך שבוע בשעת התרגול
 מרצה: שולי וינטנר. מתרגל: שלמה יונה מבוא למדעי המחשב מועד ב', סמסטר א' תשס"ג, 17/2/03 משך המבחן: שעתיים וחצי. חומר עזר: מותר כל חומר עזר, מלבד מחשב. הנחיות: ודאו כי בטופס שבידיכם 8 עמודים. יש לכתוב את
מרצה: שולי וינטנר. מתרגל: שלמה יונה מבוא למדעי המחשב מועד ב', סמסטר א' תשס"ג, 17/2/03 משך המבחן: שעתיים וחצי. חומר עזר: מותר כל חומר עזר, מלבד מחשב. הנחיות: ודאו כי בטופס שבידיכם 8 עמודים. יש לכתוב את
מבוא למדעי המחשב
 מבוא למדעי המחשב תכנות מונחה אובייקטים תוכן עניינים טיפוסי נתונים מורכבים המחלקה String תזכורת לשימוש שעשינו במחלקה String str = new String( Hello ); s.o.p(str.tostring()); s.o.p(str.charat(4)); s.o.p(str.equals(
מבוא למדעי המחשב תכנות מונחה אובייקטים תוכן עניינים טיפוסי נתונים מורכבים המחלקה String תזכורת לשימוש שעשינו במחלקה String str = new String( Hello ); s.o.p(str.tostring()); s.o.p(str.charat(4)); s.o.p(str.equals(
שבוע 4 סינטקס של HACK ASSEMBLY ניתן להשתמש בשלושה אוגרים בלבד:,A,D,M כולם בעלי 16 ביטים. M אינו אוגר ישיר- הוא מסמן את האוגר של ה RAM שאנחנו מצביעים ע
 שבוע 4 סינטקס של HACK ASSEMBLY ניתן להשתמש בשלושה אוגרים בלבד:,A,D,M כולם בעלי 16 ביטים. M אינו אוגר ישיר- הוא מסמן את האוגר של ה RAM שאנחנו מצביעים עליו כרגע )A מצביע עליו(. יש שני סוגי פקודות, פקודת
שבוע 4 סינטקס של HACK ASSEMBLY ניתן להשתמש בשלושה אוגרים בלבד:,A,D,M כולם בעלי 16 ביטים. M אינו אוגר ישיר- הוא מסמן את האוגר של ה RAM שאנחנו מצביעים עליו כרגע )A מצביע עליו(. יש שני סוגי פקודות, פקודת
(Microsoft PowerPoint - \344\370\366\340\ \372\370\356\345\353\351\356\351\344.ppt)
") תרמוכימיה כימיה פיסיקלית - 6967 האנרגיה הפנימית פנימית: אנרגיה הכוללת של המערכת האנרגיה תמיד ניתן למדוד את האנרגיה הפנימית של לא המערכת U=q+W U=U final -U initial 4 דני פורת ד"ר Tel: -6586948 e-mail: porath@chem.ch.huji.ac.il
תרמוכימיה כימיה פיסיקלית - 6967 האנרגיה הפנימית פנימית: אנרגיה הכוללת של המערכת האנרגיה תמיד ניתן למדוד את האנרגיה הפנימית של לא המערכת U=q+W U=U final -U initial 4 דני פורת ד"ר Tel: -6586948 e-mail: porath@chem.ch.huji.ac.il
שעור 6
 שעור 6 Open addressing אין רשימות מקושרות. (נניח שהאלמנטים מאוחסנים בטבלה עצמה, לחילופין קיים מצביע בהכנסה המתאימה לאלמנט אם אין שרשור). ב- addressing open הטבלה עלולה להימלא ב- factor α load תמיד. במקום
שעור 6 Open addressing אין רשימות מקושרות. (נניח שהאלמנטים מאוחסנים בטבלה עצמה, לחילופין קיים מצביע בהכנסה המתאימה לאלמנט אם אין שרשור). ב- addressing open הטבלה עלולה להימלא ב- factor α load תמיד. במקום
י( חברת פרטנר תקשורת בע"מ ("החברה" או "פרטנר") תשקיף מדף ("התשקיף" או תשקיף המדף") מכוח תשקיף מדף זה, תוכל החברה להנפיק מניות רגילות, רשומות על שם, בנ
 תשקיף מדף (התשקיף או תשקיף המדף) מכוח תשקיף מדף זה, תוכל החברה להנפיק מניות רגילות, רשומות על שם, בנ") י( חברת פרטנר תקשורת בע"מ ("החברה" או "פרטנר") תשקיף מדף ("התשקיף" או תשקיף המדף") מכוח תשקיף מדף זה, תוכל החברה להנפיק מניות רגילות, רשומות על שם, בנות 0.01 ש"ח ערך נקוב כל אחת של החברה (להלן: "המניות")
י( חברת פרטנר תקשורת בע"מ ("החברה" או "פרטנר") תשקיף מדף ("התשקיף" או תשקיף המדף") מכוח תשקיף מדף זה, תוכל החברה להנפיק מניות רגילות, רשומות על שם, בנות 0.01 ש"ח ערך נקוב כל אחת של החברה (להלן: "המניות")
התפתחות תקינה ופתולוגית
 אוניברסיטת בן-גוריון בנגב הפקולטה למדעי הרוח והחברה סילבוס קורס לסטודנט המחלקה לחינוך תשע"ז 2016-2017 התפתחות תקינה ופתולוגית - קורס בחירה 4 נק"ז מס' קורס סמסטר א': 12920011 מס' קורס סמסטר ב': 12920022
אוניברסיטת בן-גוריון בנגב הפקולטה למדעי הרוח והחברה סילבוס קורס לסטודנט המחלקה לחינוך תשע"ז 2016-2017 התפתחות תקינה ופתולוגית - קורס בחירה 4 נק"ז מס' קורס סמסטר א': 12920011 מס' קורס סמסטר ב': 12920022
Slide 1
 בית הספר למדעי המחשב אוניברסיטת תל אביב תוכנה 1 תרגול מספר 9: הורשה מחלקות אבסטרקטיות ואתחול עצמים 2 ירושה ממחלקות קיימות ראינו בהרצאה שתי דרכים לשימוש חוזר בקוד של מחלקה קיימת: הכלה + האצלה הכלה כאשר
בית הספר למדעי המחשב אוניברסיטת תל אביב תוכנה 1 תרגול מספר 9: הורשה מחלקות אבסטרקטיות ואתחול עצמים 2 ירושה ממחלקות קיימות ראינו בהרצאה שתי דרכים לשימוש חוזר בקוד של מחלקה קיימת: הכלה + האצלה הכלה כאשר
תרגול 1
 מבוא למדעי המחשב 2019 תרגול 9 חזרה על תכנות מונחה עצמים Part II ראינו בהרצאה ממשקים Set Iterable Iterator בתרגול היום חזרה על: ממשקים Iterator Iterable )iterator נראה: )set ממשק )מימוש SetAsLinkedList
מבוא למדעי המחשב 2019 תרגול 9 חזרה על תכנות מונחה עצמים Part II ראינו בהרצאה ממשקים Set Iterable Iterator בתרגול היום חזרה על: ממשקים Iterator Iterable )iterator נראה: )set ממשק )מימוש SetAsLinkedList
Thoughts from the Child Section
 מהי מטרה משמעותית? עבודת EMDR עם ילדים ברברה ויזנסקי כשאני עושה הכשרה של טיפול EMDR בילדים או מדריכה מטפל חדש בעבודתו ב- EMDR עם ילדים, אני מתחברת כל פעם מחדש להתלהבות שאני חשה בעבודה זו שטמון בה פוטנציאל
מהי מטרה משמעותית? עבודת EMDR עם ילדים ברברה ויזנסקי כשאני עושה הכשרה של טיפול EMDR בילדים או מדריכה מטפל חדש בעבודתו ב- EMDR עם ילדים, אני מתחברת כל פעם מחדש להתלהבות שאני חשה בעבודה זו שטמון בה פוטנציאל
החינוך הגופני בבית הספר מה רצוי ? מהו מקומה ש המכללה?
 חינוך גופני מתפיסת עולם לתוכנית לימודים המכללה ד"ר איתן אלדר האקדמית בוינגייט מה משפיע על תפיסת עולם של המורה? התנסות עבר )מה שהיה הוא שיהיה??( שירות צבאי תפיסה / הכתבה פוליטית גלובלית ומקומית מערכת הכשרה
חינוך גופני מתפיסת עולם לתוכנית לימודים המכללה ד"ר איתן אלדר האקדמית בוינגייט מה משפיע על תפיסת עולם של המורה? התנסות עבר )מה שהיה הוא שיהיה??( שירות צבאי תפיסה / הכתבה פוליטית גלובלית ומקומית מערכת הכשרה
PowerPoint Presentation
 תוכנה 1 תרגול 1: סביבת העבודה ומבוא ל- Java אלכסיי זגלסקי ויעל אמסטרדמר 1 בירוקרטיה אלכסיי זגלסקי שעת קבלה: שני 13:00-14:00, בתיאום מראש משרד: בניין הנדסת תוכנה, חדר 209 יעל אמסטרדמר שעת קבלה: חמישי 15:00-16:00,
תוכנה 1 תרגול 1: סביבת העבודה ומבוא ל- Java אלכסיי זגלסקי ויעל אמסטרדמר 1 בירוקרטיה אלכסיי זגלסקי שעת קבלה: שני 13:00-14:00, בתיאום מראש משרד: בניין הנדסת תוכנה, חדר 209 יעל אמסטרדמר שעת קבלה: חמישי 15:00-16:00,
Microsoft PowerPoint - 07_tdd.ppt
 המצגת מבו ס סת על הספר: Test-Driven Development By Example By Kent Beck Publisher: Addison Wesley Date: November 08, 2002 ISBN: 0-321-14653-0 Pages: 240 תכנות מונחה בדיקות (Test Driven Development) אוהד
המצגת מבו ס סת על הספר: Test-Driven Development By Example By Kent Beck Publisher: Addison Wesley Date: November 08, 2002 ISBN: 0-321-14653-0 Pages: 240 תכנות מונחה בדיקות (Test Driven Development) אוהד
מבנה מחשבים ספרתיים
 זיכרון מטמון מבנה מחשבים ספרתיים 234267 הבעיה מהירות הגישה לזיכרון איטית ביחס לביצועי המעבד )עד פי (, ככל שהזיכרון גדול יותר הגישה אליו איטית יותר. ביצועי המעבד נפגעים משמעותית אם בכל קריאה מהזיכרון יש
זיכרון מטמון מבנה מחשבים ספרתיים 234267 הבעיה מהירות הגישה לזיכרון איטית ביחס לביצועי המעבד )עד פי (, ככל שהזיכרון גדול יותר הגישה אליו איטית יותר. ביצועי המעבד נפגעים משמעותית אם בכל קריאה מהזיכרון יש
רשימת החברות החורגות
 מח' איכות הסביבה / יחידת הניטור הסביבתית Environmental Monitoring Unit Monthly Report: January דו"ח חודשי:ינואר 207 Noise Excesses Monitoring Flight Track Surveillance Noise Statistics Runway Usage Distribution
מח' איכות הסביבה / יחידת הניטור הסביבתית Environmental Monitoring Unit Monthly Report: January דו"ח חודשי:ינואר 207 Noise Excesses Monitoring Flight Track Surveillance Noise Statistics Runway Usage Distribution
הטכניון מכון טכנולוגי לישראל הפקולטה למדעי המחשב הוראות הגשה: ההגשה בזוגות. הוסיפו שמות, ת.ז., אי-מייל, תא אליו יש להחזיר את התרגיל ואת תשובותיכם לתרג
 הטכניון מכון טכנולוגי לישראל הפקולטה למדעי המחשב הוראות הגשה: ההגשה בזוגות. הוסיפו שמות, ת.ז., אי-מייל, תא אליו יש להחזיר את התרגיל ואת תשובותיכם לתרגיל, הדפיסו והגישו לתא הקורס בקומה. מבנה מחשבים ספרתיים
הטכניון מכון טכנולוגי לישראל הפקולטה למדעי המחשב הוראות הגשה: ההגשה בזוגות. הוסיפו שמות, ת.ז., אי-מייל, תא אליו יש להחזיר את התרגיל ואת תשובותיכם לתרגיל, הדפיסו והגישו לתא הקורס בקומה. מבנה מחשבים ספרתיים